Dr Rajiv Desai
An Educational Blog
NIPAH
Nipah:
_____

Figure above shows burial of a victim of Nipah virus in Kozhikode, India.
_____
Prologue:
An outbreak of probable encephalitis hit Siliguri, India in 2001. All hell broke loose when a cardiologist by the name of Dr Ajit Maity and a nurse associated with Medinova Florence Nursing Home fell victim to this disease. It was reported that both had contracted it while treating a patient. A few other doctors, nurses and paramedics were taken severely ill in another hospital. Ten among those infected medics later died. Soon, Siliguri started resembling a ghost town. The roads were deserted, shops shut and schools were closed for a week. There was misinformation circulated based on rumour and gossip. But worst of all was that some doctors secretly fled. Some doctors were apprehended at Jalpaiguri Railway Station and Bagdogra Airports by citizens before they could slip away. Red-faced politicians had flown in doctors from Kolkata. Patients were isolated immediately. Out of the 66 cases reported, 49 had died. Laboratory investigations at the time of the outbreak did not identify an infectious agent. Clinical material obtained during the Siliguri outbreak was retrospectively analyzed for evidence of Nipah virus (NiV) infection. NiV-specific immunoglobulin M (IgM) and IgG antibodies were detected in 9 of 18 patients.
At around 2 a.m. on May 17, 2018 a grievously sick Mohammed Salih, a 28-year-old architect from Kerala’s Perambra town, was rushed by his family to Kozhikode’s Baby Memorial Hospital. Salih was vomiting, had a high fever, and was in a mentally agitated state. His heart was racing at over 180 beats per minute and his blood pressure had shot up. His limbs were limp, displaying no reflexes. These symptoms were unlike any encephalitis cases that the doctors had ever seen. Nipah virus cases at Perambra in Kozhikode district of Kerala have attracted the attention of health professionals from around the world. The way Kerala has handled the Nipah virus outbreak holds crucial lessons for the rest of India. Nipah virus is an emerging virus infection that causes disease outbreaks with high case fatality rates in South-East Asia region. Nipah virus causes encephalitis and systemic vasculitis, sometimes in combination with respiratory disease. Pteropus species fruit bats are the natural reservoir of Nipah virus and zoonotic transmission can occur directly or via an intermediate host; human-to-human transmission occurs regularly. The virus was named nipah virus after Kampung Sungai Nipah (Nipah River Village in Malaysia) where the virus was isolated for the first time from pigs presenting with neurological and respiratory symptoms in 1999. Please read my article “Ebola” published on December 6, 2014 on this website as both Ebola and Nipah are deadly zoonotic viruses transmitted from bats to humans.
______
Abbreviations and synonyms:
Nipah virus disease = Nipah = Barking Pig Syndrome = Porcine Respiratory and Encephalitis Syndrome
NiV = nipah virus
NIV = national institute of virology, India
NiV-B = NiV Bangladesh strain = bNiV = NiV-BD
NiV-M = NiV Malaysia strain = mNiV = NiV-MY
Fruit bat = flying fox = bats of the genus Pteropus = pteropid bats
R0 = basic reproduction number
HeV = hendra virus
BSL-4 = biosafety level 4
JE = Japanese encephalitis
RT-PCR = reverse transcription-polymerase chain reaction
TCID50 = 50% Tissue Culture Infective Dose
PPE = personal protective equipment
PFU = plaque-forming unit
______
______
Nipah (NiV) and Hendra (HeV) Viruses:
Nipah virus (NiV) and Hendra virus (HeV) are emerging zoonotic viruses that can be transmitted from bats to humans directly or via intermediary hosts. Nipah and Hendra viruses are two zoonotic paramyxoviruses with an ability to cause fatal encephalitic and respiratory diseases in humans. These are the only two viruses in the Paramyxoviridae family (genus henipavirus) classified as biosafety level 4 (BSL4), and their mortality rates in humans are 40 to 100%. Hendra virus was first identified in humans in Australia in 1994, with horses as the intermediate host. HeV is transmitted to horses via fruit bats, and several outbreaks in horses have been reported as recently as 2014. Nipah virus emerged in humans in 1998 in Malaysia, with pigs as the intermediate host. Nipah virus was later also identified in India and Bangladesh in 2001, with the flying fox (Pteropus spp.) as the natural host, although no intermediary animal host was found in more recent outbreaks there. A third zoonotic paramyxovirus, Menangle virus, was first identified in pigs. Of the zoonotic paramyxoviruses, Nipah virus is responsible for the greatest number of human cases, with several hundred cases and at least 365 deaths reported, compared to Hendra virus, which has caused a handful of cases and 2 deaths, and Menangle, which has only caused self-limited illness in 2 individuals.
______
Emerging diseases:
Diseases that are rapidly increasing in incidence or distribution are said to be ‘emerging’. The definition encompasses not only diseases associated with previously unknown (or novel) agents, but also those known diseases that are ‘re-emerging’ spatially or temporally. What triggers disease emergence? Modern epidemiological principles contend that disease is multi-factorial – that in addition to the presence of the infectious agent, additional factors are generally necessary for infection and disease to occur. Such factors may relate to the agent, to the host, or to the environment. Putative contributing factors to disease emergence include ecological changes, changes in human demographics and behaviour, increased international travel and commerce, advances in technology and industry, microbial adaptation or change, and breakdown of public health measures (Morse 1995).
Many emerging infections are zoonoses. The introduction of a “new” zoonotic infection into a human or domestic animal population can follow the incursion of humans (accompanied by their domestic animals, livestock, and crops) into previously remote natural habitats where unknown disease agents exist in harmony with wild reservoir hosts. Upon contact with new and naive species, an agent may ‘jump species’ and establish in a new species which has no natural immunity or evolved resistance (unlike the natural host which may have evolved with the agent over time). The maintenance of monocultures of genetically similar or identical individuals may further promote susceptibility to infection. Further, artificially maintained high population densities may facilitate the rapid spread of pathogens throughout livestock populations. Zoonotic infections may be passed directly to humans from the natural reservoir, or they may be transmitted to humans via an intermediate, amplifying host.
______
______
Zoonotic diseases:
According to the World Health Organization (WHO), “A zoonosis is any disease transmitted from vertebrate animals to humans.” Throughout the history of the world humans have been plagued by diseases of various types and origins. More than 60% of the newly identified infectious agents that have affected people over the past few decades have been caused by pathogens originating from animals or animal products. Of these zoonotic infections, 70% originate from wildlife. Zoonotic diseases, or diseases which have the capability to jump species, animals to humans or vice versa, have been particularly troublesome and deadly. Zoonotic diseases are unique in that they are mainly caused by pathogens such as fungi, bacteria, parasites, or viruses. These pathogens typically survive in a reservoir host, which have immunity to the pathogen. The list of possible reservoir hosts capable of transmitting disease to humans is expansive; however the most common are apes, insects, rodents, and bats. The diseases are then passed to humans who come in contact with an infected animal through bites or scratches, an infected animal’s environment, or animal secretions such as saliva, feces, or mucus. Often these diseases have a higher virulence because of the lack of any immunity within the human population and the ease of transmission. Some examples of zoonotic diseases include anthrax, bird flu, ebola, dengue, rabies, malaria, swine flu and leptospirosis. As of recent, more and more zoonotic diseases are emerging because of an increase in human and wildlife interaction. An increase in farming and deforestation has resulted in humans and wildlife into the same habitat. A prime example of this is the emergence of the Nipah virus (NiV). NiV is a member of the Henipavirus genus in the Paramyxoviridae, and has become a growing concern of Southeast Asia and Australia. Nipah virus is also a growing concern for the United States. The Center of Disease Control (CDC) has declared it a biosafety level 4 agent. This the highest biosafety level category, home to agents which can be distributed via aerosol transmission and have no treatment or vaccine. Similar biosafety level 4 agents are Ebola, Smallpox, and several hemorrhagic diseases. The CDC has also tagged the Nipah virus a Category C bioagent, the third highest priority agent category in regards to biological warfare. The availability, simplicity to produce and disperse, and high mortality rate of the Nipah virus make it possible for it to be used as a weapon of biological warfare.
_
Over the past decade, the previously unknown paramyxoviruses Hendra virus (HeV) and Nipah virus (NiV) have emerged in humans and livestock in Australia and Southeast Asia during the 1990s. Like the avian flu, SARS, and Ebola viruses, Hendra and Nipah are zoonotic pathogens. That means they originate in certain animals but can jump between animal species and between animals and humans. Harbored by fruit bats, they cause potentially fatal encephalitis and respiratory disease in humans, with a devastating 75 percent fatality rate. Both viruses are contagious, highly virulent, and capable of infecting a number of mammalian species and causing potentially fatal disease. Due to the lack of a licensed vaccine or antiviral therapies, HeV and NiV are designated as biosafety level (BSL) 4 agents and are potential bioterrorist agents. The genomic structure of both viruses is that of a typical paramyxovirus. However, due to limited sequence homology and little immunological cross-reactivity with other paramyxoviruses, HeV and NiV have been classified into a new genus within the family Paramyxoviridae named Henipavirus. HeV emerged in Queensland, Australia, in 1994, killing 1 human and 14 horses, and it was responsible for at least 4 other sporadic outbreaks involving horses and humans between 1994 and 2006. The closely related NiV emerged in 1998–1999 in peninsular Malaysia, resulting in the death of >100 people and the culling of >1 million pigs. Since then, several NiV outbreaks have been recorded in Bangladesh and India. Nipah virus (NiV) is a paramyxovirus whose reservoir host is fruit bats of the genus Pteropus. Occasionally the virus is introduced into human populations and causes severe illness characterized by encephalitis or respiratory disease.
______
______
Bats:
Bats are mammals of the order Chiroptera; with their forelimbs adapted as wings. Bats are more manoeuvrable than birds, flying with their very long spread-out digits covered with a thin membrane or patagium. Bats are the only mammals capable of sustained flight, as opposed to gliding, as in the flying squirrel. The fastest bat, the Mexican free-tailed bat (Tadarida brasiliensis), can achieve a ground speed of 160 kilometres per hour (99 mph). Seasonal movements from hibernacula and/or swarms to summer colonies ranged widely from 10 to 647 km. The second largest order of mammals, bats comprise about 20% of all classified mammal species worldwide, with over 1,200 species. Many bats are insectivores, and most of the rest are frugivores (fruit-eaters). A few species feed on animals other than insects; for example, the vampire bats feed on blood. Most bats are nocturnal, and many roost in caves or other refuges; it is uncertain whether bats have these behaviours to escape predators. Bats are present throughout the world, with the exception of extremely cold regions. They are important in their ecosystems for pollinating flowers and dispersing seeds; many tropical plants depend entirely on bats for these services. They are natural reservoirs of many pathogens, such as rabies; and since they are highly mobile, social, and long-lived, they can readily spread disease. The maximum lifespan of bats is three-and-a-half times longer than other mammals of similar size. Five species have been recorded to live over 30 years in the wild. In many cultures, bats are popularly associated with darkness, malevolence, witchcraft, vampires, and death. Nipah Virus (NiV) represents another new emerging zoonosis, one of the most important bat-borne pathogens discovered in recent history. Nipah is believed to be transmitted from what are called flying foxes, or mega bats, so called because they are the largest bat species. They eat fruits and live in trees. These are a part of the old-world fruit bat family, called pteropid bats. Having been around for millions of years, bats have probably carried infectious diseases for nearly as long. Bats often end up being reservoirs for a number of severe infectious diseases, including Ebola, SARS coronavirus, MERS corona virus, Marburg, Melaka, Nipah and Hendra. Several bat species can carry these viruses that are deadly to humans without getting sick themselves. But scientists say that villainizing bats is not the answer. They’re a crucial part of their ecosystems. They are also really important pollinators. Several factors have increased the chance of bat-borne viruses being passed humans, including development that has encroached on the bats’ natural habitats.
_
What makes the bat hospitable to so many viruses?
Bats host many more zoonotic viruses than their closest competitors, rodents. To qualify as a natural reservoir for a virus, an animal should harbour high virus diversity, live in large gregarious groups, travel widely, and live long enough for adequate dispersal. This last requirement is defeated if the virus makes the animal sick enough to die. Bats offer all household conveniences for viruses to replicate, diversify, and disperse widely across many host species. Bats are social creatures and aggregate in large groups. Caste and creed are no bar as many species roost together. So there is active viral exchange, and with each exchange, viral diversification allows novel forms to emerge. Bats support replication and circulation of high titers of virus without becoming ill. A perfect parasite is able to replicate and not kill its host which is an evolutionary advantage to remain endemic in its host species population. All bats can carry viruses, some of them deadly. The Marburg virus, a relative of Ebola, was isolated in 2009 from the Egyptian Rousette, a fruit bat, in Uganda’s Kitaka Cave. After the 2003 outbreak of Severe Acute Respiratory Syndrome (SARS) in China, researchers found antibodies to the SARS Coronavirus in cave-dwelling insectivorous bats. Similarly, Ebola antibodies were found in species like the Hammer-headed fruit bat. Why are so many emerging diseases linked to bats? For one thing, with around 1,200 species, bats comprise 20% of the earth’s mammalian diversity. So, it ought not to be surprising that they host many viruses. Not all of these viruses are threats to humans. The bigger question is how bats stay healthy despite carrying these pathogens. The Indian Flying Fox, for example, hosts over 50 viruses. So far, researchers have only hypotheses to explain this viral diversity in bats. One explanation — the “flight as fever” hypothesis — suggests that long periods of flying raises the temperatures of bats, boosting their immune responses. This helps them survive the microbes’ pathogenic effects.
______
______
Introduction to nipah virus (NiV) disease:
_
Nipah virus (NiV) is a newly emergent zoonotic pathogen of the family Paramyxoviridae that can cause rapid, fatal respiratory and neurologic disease in both humans and animals (Chua et al., 2000; Wong et al., 2002; Bishop and Broder, 2008). Nipah is a viral zoonotic disease caused by NiV of the Henipavirus genus of Paramyxoviridae family. Pteropus bats (fruit eating species, popularly known as flying foxes) are supposed to be the natural hosts of the virus. Nipah virus is similar to Hendra virus that was discovered in Australia in 1994. Nipah virus was discovered in 1999. Nipah and Hendra viruses are two related zoonotic pathogens that have emerged in the Asia-Pacific region. Both are RNA viruses belonging to the Paramyxoviridae family and grouped under the genus Henipavirus, since they share antigenic, serological, and ultrastructural characteristics and differ from other paramyxoviruses. Another virus in the genus is the non-pathogenic Cedar virus. Nipah virus caused an outbreak in pigs and humans in Malaysia and Singapore between 1998 and 1999, and has caused recurrent human outbreaks in Bangladesh and West Bengal, India since 2001. Latest nipah outbreak was reported in India’s Kerala state in May 2018. An outbreak of henipavirus infection, most likely due to Nipah or a Nipah-like virus, occurred in the Philippines in 2014. Hendra virus infections affecting horses and humans have occurred in Australia since 1994. Given the relatedness of NiV to Hendra virus, bat species were quickly singled out for investigation and flying foxes of the genus Pteropus were subsequently identified as the reservoir for NiV although evidence of henipavirus infection has now been reported in a wider range of both frugivorous and insectivorous bats. Nipah cases tend to occur in a cluster or as an outbreak.
__
Hendra virus (HeV) first appeared in Australia in 1994, with infection and fatal disease occurring in horses and humans. In total, two of three infected horse handlers and 15 horses succumbed to the fatal HeV disease. Nipah virus (NiV) appeared in peninsular Malaysia in 1998 in pigs and pig farmers. By mid-1999, more than 265 human cases of encephalitis, including 105 deaths, had been reported in Malaysia, and 11 cases of either encephalitis or respiratory illness with one fatality were reported in Singapore. Although HeV and NiV emerged independently, further characterization demonstrated that both viruses were paramyxoviruses that have similar biological, molecular, and serological properties that were distinct from those of all other paramyxoviruses, and consequently, they were grouped together as closely related viruses in the new Henipavirus genus. The known natural reservoir hosts of both HeV and NiV are pteropid fruit bats, commonly known as flying foxes, which do not exhibit clinical disease when infected. Numerous flying fox species have antibodies to HeV and NiV, and their vast geological range overlaps with all henipavirus outbreaks. Unlike all other paramyxoviruses, HeV and NiV have a broad species tropism, and in addition to infecting bats, they can infect and cause disease, often with very high fatality rates, in a wide range of species spanning six mammalian orders.
_
Nipah virus (NiV) is a paramyxovirus in the genus Henipavirus. Two lineages are currently circulating in Southeast Asia: a Malaysian strain (mNiV) and a Bangladesh strain (bNiV). Experimentally, mNiV appears to be the more virulent of the lineages. NiV is closely related to Hendra virus (HeV), which causes high mortality in both horses and humans and is endemic in parts of Australia. The best known reservoir for NiV, the flying fox, is confined to Southeast Asia, Australia, and eastern Africa. NiV can infect many different species, including bats, humans, cats, dogs, pigs, and other livestock. Pigs are an amplifying host for NiV and person-to-person transmission can also occur. NiV-infected humans present with encephalitis, and case fatality rates can reach 75%. Workers with direct contact with infected swine are most at risk for contracting NiV during an outbreak. Pigs transmit NiV to people through sputum, splashing urine, and large respiratory droplets, facilitated by a characteristic barking cough in infected pigs. Transmission between pigs occurs through direct contact, exposure to infected secretions, and contact droplet transmission.
_
In a 2005 review on emerging and reemerging infectious agents, of the 1407 human pathogens, 816 (58%) were classified as zoonotic in origin. In recent decades, zoonotic pathogens have induced considerable stress and anxiety in a broad range of societies worldwide. The emergence of Nipah virus (NiV) in Peninsular Malaysia in September 1998 was the second in a series of spillover events. The first, starting in September 1994, was an outbreak of Hendra virus (HeV) in Brisbane, Australia. Nipah and Hendra viruses are members of the family Paramyxoviridae (genus: Henipavirus), each can potentially cause fatal disease in human and animal hosts. In Malaysia, Nipah was first detected in 1999. Several people who were engaged in rearing pigs at Sungai Nipah village on the banks of Nipah River, fell ill and died. When authorities conducted an investigation, they found that the disease was contracted from pigs. The pigs were infected with the virus after they consumed the fruits partially eaten by bats. Authorities noticed that migratory bats had spread the disease. The fruits that the pigs ate had traces of saliva of the bats. People who had touched the secretions from the nose and other areas of pigs fell sick. The virus had spread from animal to man, but not from man to man. NiV-infected pigs developed a unique clinical condition called ‘barking pig syndrome’. The first human cases in Malaysia (Perak, Negri Sembilan, and Selangor states) and Singapore were reported amongst abattoir workers. The Malaysia epidemic resulted in 265 cases of acute encephalitis with 105 deaths and the culling of 1.1 million pigs. Since 1998, Malaysia and Singapore have not documented human cases; however, human disease has been continuously reported in Bangladesh since 2001, with mortality rates estimated at approximately 70%. Subsequently, NiV has emerged as a significant public health threat in Bangladesh and India. Unlike the initial outbreak, in which pigs were the intermediate host, the role of bat reservoirs in human infection has been substantiated. The geography of NiV in Bangladesh, exhibits characteristics of clustering, particularly in the Dhaka, Khulna, Rajshahi, and Rangpur divisions. Date palm monoculture and the geographic distribution of transmission events since 2001 display strong spatial dependency. A number of cases have been linked to drinking raw date palm sap, which had probably been contaminated by bats. Drinking fermented date palm sap (alcohol content approximately 4%) appeared to be a risk factor in a few cases. The drinking of raw date palm sap contaminated with fruit bat urine or saliva containing NiV is the only known cause of outbreak of the disease in Bangladesh outbreaks. Bats (order: Chiroptera) of the family Pteropodidae, genus Pteropus (flying foxes) are the presumed wildlife reservoir of NiV. Pteropus giganteus or the Indian flying fox is the largest frugivorous bat species in Bangladesh and is of key interest as the zoonotic reservoir of Nipah virus. Pteropus giganteus is further associated with harbouring at least 55 recently-discovered viruses. The asymptomatic nature of NiV in bats suggests that the virus had evolved alongside Pteropus bats for centuries, and more than likely this adaptation has been responsible for human exposure long before the virus was first reported in 1998. Biological traits making bats well-suited for hosting a variety of microorganisms include their long lifespans, which facilitate viral persistence and their ability for flight. Long-distance travel is prevalent; in fact, the grey-headed flying fox (Pteropus poliocephalus) expands its range by up to 600 km during long-distance travel between roosting sites. Regionally, 330 species of bats are endemic to Southeast Asia, which accounts for 25% of the world’s overall bat diversity. The genus Pteropus alone features 60 species of bat with broad geographic distributions extending to the east coast of Africa, the Philippines, Indonesia, New Guinea, and much of the Indian sub-continent.
_
The emergence of NiV into the pig population and subsequently into the human population is believed to be due to changes in ecological conditions. Urbanization, deforestation and drought resulting in a shortage of resources for bat populations could have compelled bats to move from their natural habitats to agricultural areas. Among the factors that contributed to the disease emergence in Malaysia is the establishment of pig farms within the range of natural host that led to the initial introduction into the pig population; the maintenance of high densities of pigs led to the rapid dissemination of the infection within local pig populations; and the transport of pigs to other geographic areas for commerce led to the rapid spread of disease in pigs in southern Malaysia and Singapore. The presence of high density, amplifying host population facilitated transmission of the virus to human. The same may be true for the NiV outbreaks in Bangladesh and India, and here it is postulated that the outbreak in humans may be due to direct contact with bats or indirectly by contact with material contaminated by bats. It is apparent from the presence of the virus and antibodies in the fruit bats of the region and 13 years of continuous NiV outbreaks in humans in Bangladesh that it is the potential threat to the Indian subcontinent. The medical and veterinary professionals should also increase the awareness of the disease particularly hosts and mode of transmission of the virus.
__
Nipah virus infection (NiV) is a viral infection caused by the Nipah virus. Nipah virus infection in humans causes a range of clinical presentations, from asymptomatic infection (subclinical) to acute respiratory infection and fatal encephalitis. Symptoms from infection vary from none to fever, cough, headache, shortness of breath, and confusion. This may worsen into a coma over a day or two. Complications can include inflammation of the brain and seizures following recovery. The case fatality rate is estimated at 40% to 75%. This rate can vary by outbreak depending on local capabilities for epidemiological surveillance and clinical management. The Nipah virus is a type of RNA virus in the genus Henipavirus. It can both spread between people and from other animals to people. Nipah virus can be transmitted to humans from animals (such as bats or pigs), or contaminated foods and can also be transmitted directly from human-to-human. Spread typically requires direct contact with an infected source. The virus normally circulates among specific types of fruit bats. Fruit bats of the Pteropodidae family are the natural host of Nipah virus. Diagnosis is based on symptoms and confirmed by laboratory testing. Management involves supportive care. As of 2018 there is no vaccine or specific treatment. Prevention is by avoiding exposure to bats and sick pigs and not drinking raw date palm sap. As of May 2018 about 608 human cases of Nipah virus are estimated to have occurred and 50 to 75 percent of those who were infected died. In May 2018, an outbreak of the disease resulted in at least 17 deaths in the Indian state of Kerala. Although Nipah virus has caused only a few known outbreaks in Asia, it infects a wide range of animals and causes severe disease and death in people, making it a public health concern.
_
Henipavirus’s broad species tropism and ability to cause fatal respiratory and/or neurologic disease in humans and animals make them important transboundary biological threats. Over the past decade a considerable amount of research has focused on the henipavirus envelope glycoproteins and their roles in the virus attachment and infection process. These efforts have now led to the development and testing of both passive and active immunization strategies applicable to both human and animal use. Presently, a cross-reactive human mAb (m102.4) has been demonstrated as an exceptionally efficacious post-exposure therapy in protecting both ferrets and nonhuman primates from lethal henipavirus disease, and its effectiveness led to its application in people as a compassionate use post-exposure prophylaxis in Australia. Also, as an active vaccination strategy for preventing Hendra virus infection and disease in horses in Australia and thus blocking potential transmission to people, a recombinant subunit vaccine, HeV-sG, which has been shown to provide protection against henipavirus challenge in cats, ferrets, monkeys and now horses, has been licensed and deployed for use in Australia. To date, henipavirus antivirals have only been deployed in Australia in the fight against Hendra virus. As Nipah virus causes significantly more instances of human disease, increased efforts are needed to advance Nipah-targeted countermeasures in endemic regions. Animal models have demonstrated that both the HeV-sG vaccine and the m102.4 human antibody can prevent both Nipah virus infection and disease. Efforts are currently under way to develop HeV-sG for human use as well as for use in pigs.
____
____
According to WHO:
Nipah virus (NiV) is an emerging pathogen first identified in 1999 in Malaysia, with cases also seen in Singapore, in an outbreak of acute encephalitis in pigs and humans. Since then, human NiV outbreaks have been reported in India and Bangladesh. While no new outbreaks have been reported in Malaysia and Singapore, repeated outbreaks have been noted in Bangladesh almost every year since 2001 in select districts with occasional outbreaks in neighbouring India. From 1998 to 2015, there have been at least 600 cases of NiV human infections, with case fatalities in later outbreaks in India and Bangladesh ranging between 43 and 100%. This is a small number considering the world statistics for other diseases. Human to human transmission is particularly notable in the outbreaks in India and Bangladesh, accounting for 75% and 51% of cases, respectively. NiV infection has both a neurological and respiratory disease presentation. Respiratory involvement differs in prevalence between the outbreak in Malaysia (29%) and Bangladesh (75%). Relapsing NiV encephalitis distinct from acute NiV encephalitis has been described and is estimated to occur in <10% of survivors. NiV is related to Hendra virus (HeV), another paramyxovirus which has been classified as a member of the genus Henipavirus. Pigs and horses have been implicated as potential multiplier hosts for NiV and HeV, respectively. The primary reservoir of NiV is fruit bats of the genus Pteropus. Pigs were the intermediate hosts in the outbreaks in Malaysia and Singapore, while in Bangladesh humans were infected as a result of consuming date palm sap that had been contaminated by infected fruit bats. There is currently a licensed horse vaccine for HeV.
_
As per the fact sheet about Nipah Virus by the World Health Organization:
- Infected bats shed virus in their excretion and secretion such as saliva, urine, semen and excreta but they are symptomless carriers.
- The NiV is highly contagious among pigs, spread by coughing.
- Direct contact with infected pigs was identified as the predominant mode of transmission in humans when it was first recognized in a large outbreak in Malaysia in 1999.
- Drinking of fresh date palm sap, possibly contaminated by fruit bats (P. giganteus) during the winter season, may have been responsible for indirect transmission of Nipah Virus to humans.
- During the Bangladesh outbreak the virus is suggested to have been transmitted either directly or indirectly from infected bats to humans.
- There is circumstantial evidence of human-to-human transmission in India in 2001. During the outbreak in Siliguri, 33 health workers and hospital visitors became ill after exposure to patients hospitalized with Nipah Virus illness, suggesting nosocomial infection.
- Human-to-human transmission of NiV has been reported in recent outbreaks demonstrating a risk of transmission of the virus from infected patients to healthcare workers through contact with infected secretions, excretions, blood or tissues.
_______
_______
Classification, structure, and virology of nipah and related viruses:
_
Single stranded RNA viruses are classified as positive or negative depending on the sense or polarity of the RNA. A negative-sense single-stranded RNA virus (-ssRNA virus) is a virus that uses negative sense, single-stranded RNA as its genetic material which does not encode mRNA (messenger RNA). The negative viral RNA is complementary to the mRNA and must be converted to a positive RNA by RNA polymerase before translation. The positive-sense RNA acts as a viral mRNA, which is translated into proteins for the production of new virion materials. Therefore, the purified RNA of a negative sense virus is not infectious by itself, as it needs to be converted to a positive sense RNA for replication. Examples of negative-strand RNA viruses include influenza virus, measles viruses, and rabies virus.
_
Nipah virus is a member of the genus Henipavirus in the family Paramyxoviridae. This genus also includes Hendra virus, Cedar virus (an apparently nonpathogenic virus found in Australian bats) and additional uncharacterized henipaviruses in various locations. There seem to be multiple strains of Nipah virus. There are two lineages of NiV circulating in Southeast Asia: Malaysian NiV (mNiV) and Bangladesh NiV (bNiV). In the 1998 Malaysian NiV outbreak, only a single mNiV isolate was identified; however, an outbreak of bNiV may be caused by multiple strains. In vitro and in vivo hamster models show that mNiV causes increased cytopathology, increased disease progression, and higher mortality rates than bNiV. However, bNiV appears to cause more respiratory disease in people, leading to non-productive cough in 62% of patients vs. only 14% of patients infected with mNiV.
_
The nucleotide sequences of Nipah virus strains isolated from pigs and persons in Malaysia were remarkably similar and suggest that the entire outbreak was caused by 1 or 2 closely related strains. Indeed, all human cases of Nipah infection in Malaysia and Singapore could have originated from a single or perhaps 2 introductions of Nipah virus from its bat reservoir into pigs. In Bangladesh, by contrast, recurrent Nipah outbreaks have been recognized since 2001, and the strains of Nipah isolates show substantial heterogeneity in their nucleotide sequences. This heterogeneity suggests repeated introductions of Nipah virus from its host reservoir into the human population in Bangladesh. In 2012, Yadav et al. have surveyed the Indian states of Maharashtra and West Bengal to evaluate the presence of viral RNA and IgG against NiV in different bat populations belonging to the species Pteropus giganteus, Cynopterus sphinx and Megaderma lyra. Authors found NiV RNA in Pteropus bat thus suggesting it may be a reservoir for NiV in India. Furthermore, the phylogenetic analysis demonstrated that two phylogenetic lineages were formed for NiV sequences, one including Bangladesh and India sequences and the other Malaysia and Cambodia sequences. By phylogenetic analysis it was unmistakable confirmed that the same NiV strain circulates in India and Bangladesh and that it was different from that circulating in Malaysia and Cambodia.
_
The Paramyxoviridae family of lipid-enveloped negative-strand RNA viruses comprises several genera that include such human pathogens as the human parainfluenza virus, the measles and mumps viruses, the Newcastle disease virus (NDV), and the human respiratory syncytial virus, among others. Nipah and Hendra viruses are negative-sense, single-stranded RNA viruses in the Paramyxoviridae family, subfamily Paramyxovirinae. They are further categorized in the recently named genus Henipavirus, one of five genera in the subfamily (the others are Respirovirus, Morbillovirus, Avulavirus, and Rubulavirus) (Figure below). Other human pathogenic viruses exist in these other genera, such as measles, mumps, and parainfluenza viruses; Nipah and Hendra viruses in the genus Henipavirus, and Menangle virus in the genus Rubalavirus, are unique in that they are zoonotic and are viruses that have recently emerged in humans.
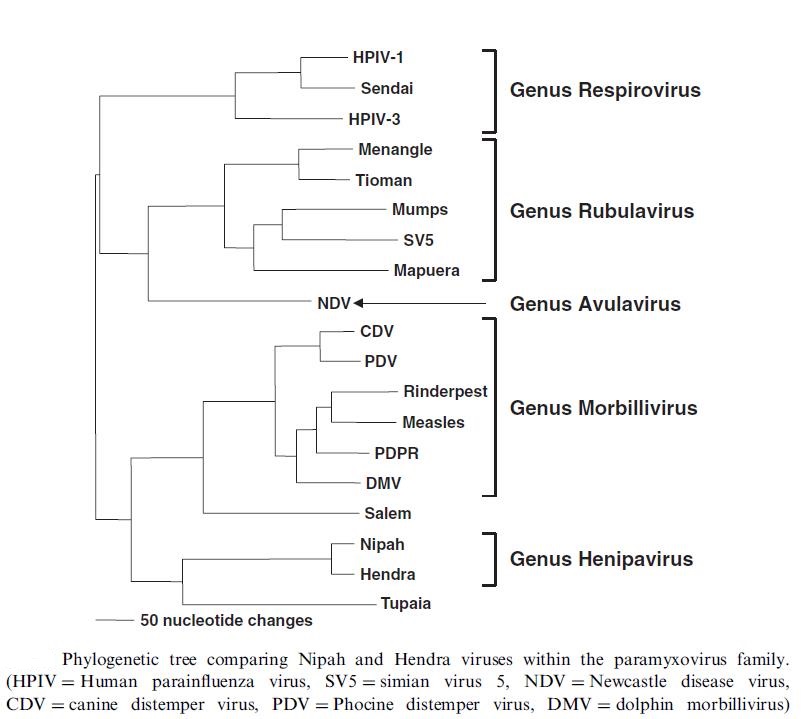
_
HeV and NiV are novel members of the family Paramyxoviridae. Paramyxoviruses are negative-sense RNA enveloped viruses and contain 2 major membrane-anchored envelope glycoproteins that are required for infection of a receptive host cell. The broad species tropisms and the ability to cause fatal disease in both animals and humans distinguish HeV and NiV from all other known paramyxoviruses. The substantial differences in their genome sequence and host range led to the establishment of a new genus (Henipavirus) in the family to accommodate their taxonomic classification. Fruit bats in the genus Pteropus (flying foxes) are the natural reservoir of both HeV and NiV, and NiV is present in fruit bat populations in Indonesia, Thailand, Malaysia, Bangladesh and Cambodia.
_
Genus Henipavirus:
Etymology: Henipavirus: from Hendra virus (Hendra after the Brisbane suburb from which it was first isolated in 1994) and Nipah virus.
_
Taxonomy of henipavirus:
Group V: Negative sense ssRNA viruses
Order: Mononegavirales
Family: Paramyxoviridae
Genus: Henipavirus
_
Phylogeny of the Mononegavirales order:
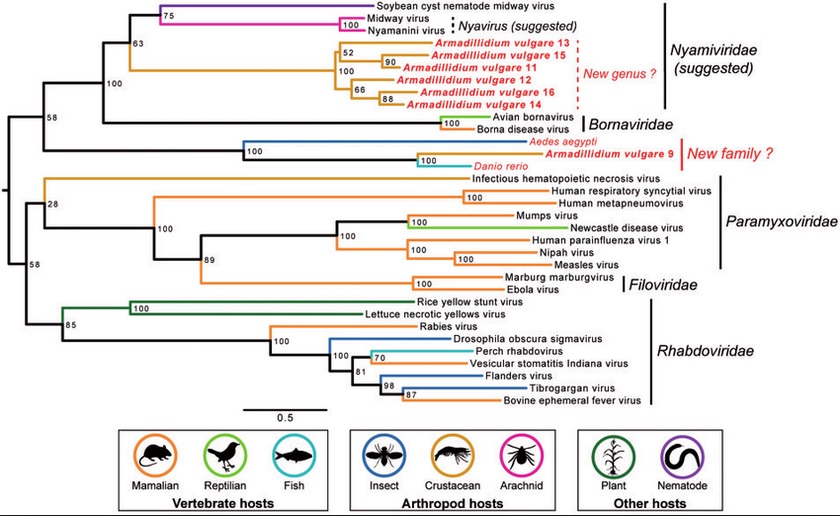
The tree was obtained from ML analysis of the RNA-dependent RNA polymerase multiple alignment, including Armadillidium vulgare EVE sequences, sequences of closely related exogenous and endogenous viruses, and representative virus species of the Mononegavirales order. ML nonparametric bootstrap values (100 replicates) are indicated at each node. Associated hosts are indicated by branch colors and silhouettes at the bottom.
_
Henipavirus is a genus of RNA viruses in the family Paramyxoviridae, order Mononegavirales containing five established species. Henipaviruses are naturally harboured by pteropid fruit bats (flying foxes) and microbats of several species. Henipaviruses are characterised by long genomes and a wide host range. Their recent emergence as zoonotic pathogens capable of causing illness and death in domestic animals and humans is a cause of concern. In 2009, RNA sequences of three novel viruses in phylogenetic relationship to known henipaviruses were detected in African straw-colored fruit bats (Eidolon helvum) in Ghana. The finding of these novel henipaviruses outside Australia and Asia indicates that the region of potential endemicity of henipaviruses may be worldwide. These African henipaviruses are slowly being characterised.
_
| Genus Henipavirus: species and their viruses | ||
| Genus | Species | Virus (Abbreviation) |
| Henipavirus | Cedar henipavirus | Cedar virus (CedV) |
| Ghanaian bat henipavirus | Kumasi virus (KV) | |
| Hendra henipavirus* | Hendra virus (HeV) | |
| Mojiang henipavirus | Mòjiāng virus (MojV) | |
| Nipah henipavirus | Nipah virus (NiV) | |
* denotes type species.
_
Henipavirus structure:
Henipavirions are pleomorphic (variably shaped), ranging in size from 40 to 600 nm in diameter. They possess a lipid membrane overlying a shell of viral matrix protein. At the core is a single helical strand of genomic RNA tightly bound to N (nucleocapsid) protein and associated with the L (large) and P (phosphoprotein) proteins, which provide RNA polymerase activity during replication. Embedded within the lipid membrane are spikes of F (fusion) protein trimers and G (attachment) protein tetramers. The function of the G protein is to attach the virus to the surface of a host cell via EFNB2, a highly conserved protein present in many mammals. The structure of the attachment glycoprotein has been determined by X-ray crystallography. The F protein fuses the viral membrane with the host cell membrane, releasing the virion contents into the cell. It also causes infected cells to fuse with neighbouring cells to form large, multinucleated syncytia.
_
General structure of a Henipavirus:
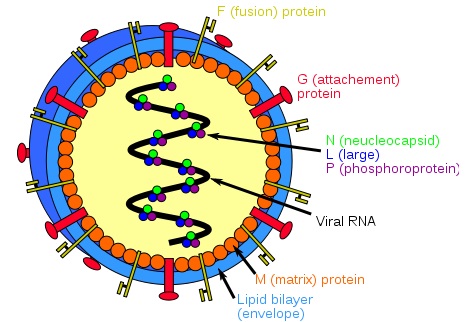
The six key proteins, P, N, F, G, M, and L, are shown in their natural position and labelled. In the center of the virion is the negative sense single stranded RNA.
_
Genome structure:
As all mononegaviral genomes, Hendra virus and Nipah virus genomes are non-segmented, single-stranded negative-sense RNA. Both genomes are 18.2 kb in length and contain six genes corresponding to six structural proteins. In common with other members of the Paramyxoviridae family, the number of nucleotides in the henipavirus genome is a multiple of six, consistent with what is known as the ‘rule of six’. Deviation from the rule of six, through mutation or incomplete genome synthesis, leads to inefficient viral replication, probably due to structural constraints imposed by the binding between the RNA and the N protein. Henipaviruses employ an unusual process called RNA editing to generate multiple proteins from a single gene. The specific process in henipaviruses involves the insertion of extra guanosine residues into the P gene mRNA prior to translation. The number of residues added determines whether the P, V or W proteins are synthesised. The functions of the V and W proteins involve disrupting host antiviral mechanisms.
______
Nipah virus:
_
Nipah virus structure:
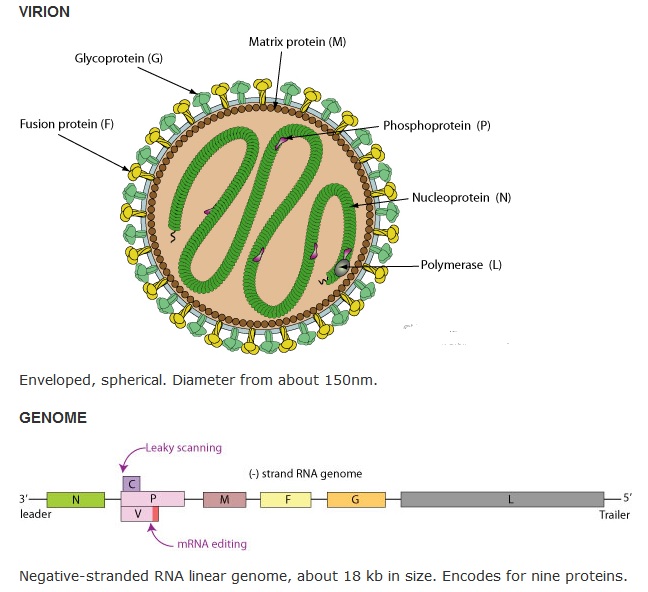
_
The NiV genome is a negative-sense nonsegmented RNA, of 18,246 nucleotides (Malaysian isolate) or 18,252 nucleotidest (Bangladesh isolate) (Harcourt et al., 2005, 2001). [18,234 nucleotides for HeV] Like other paramyxoviruses, the genome contains six transcriptional units encoding six structural proteins: nucleocapsid (N), phosphoprotein (P), matrix protein (M), fusion protein (F), receptor-binding (G), and RNA-dependent RNA polymerase (L), and three nonstructural proteins, C, V, and W (Harcourt et al., 2000). The NiV C protein (NiV-C) is expressed from an alternative open reading frame at the 5′ end of the P gene and shares no sequence similarity with the P protein, whereas the V and W proteins share the same N-terminal domain with the P protein, because they are translated from the edited P mRNA when one or two nontemplate G residues are inserted into the highly conserved RNA editing site of the P gene. It has been reported that each NiV P gene product, particularly proteins V, W, and C, has interferon (IFN)-antagonist activity (Park et al., 2003; Shaw et al., 2004). The host IFN responses induced by viral infection are involved in the early innate immune response and in the modulation of the subsequent acquired immune response. The common N-terminal domain of the NiV V and W proteins has IFN-antagonist activity (Park et al., 2003). The V protein inhibits the IFN-induced nuclear translocation and phosphorylation of signal transducer and activator of transcription (STAT) (Rodriguez et al., 2002; Shaw et al., 2004). The W protein binds STAT1 after it enters the nucleus and prevents it shuttling back to the cytoplasm (Shaw et al., 2004). In this way, the differential localization of V and W in the cytoplasm and nucleus determines the IFN-antagonist mechanism of each protein. The NiV V and W proteins also inhibit IFN synthesis (Shaw et al., 2005). The V proteins of a number of paramyxoviruses, including NiV, interact with the helicase domain of MDA5, inhibiting its ATPase activity (Childs et al., 2009; Motz et al., 2013; Rodriguez and Horvath, 2013). Unlike the V and W proteins, the mechanism of the IFN-antagonist activity of NiV-C is unknown. The C protein appears to regulate viral RNA synthesis and may play a role as a virulence factor.
_
The virion is enveloped by a traditional lipid bilayer but “spiked” with fusion (F) and receptor-binding glycoproteins (G). The fusion proteins are responsible for fusing the viral membrane to the host membrane triggering the release of the contents of the virion. The receptor-binding glycoporteins are extremely specific and bind only to Ephrin B2 (EFNB2) surface proteins. Specifically, NiV has been found to alternatively bind to EFB3 as well. The EFNB2 surface proteins are highly conserved across the mammalian lineage. On the underside of the lipid bilayer matrix proteins (M) are present for structural support and regulating the budding process. Other proteins, C, V, and W, are also present in the cytoplasm and involved in regulation of transcription and replication. In regards to the Nipah virus genome, the exact structure is not completely understood. However because of the strong homology between Hendra virus and Nipah virus, a nearly identical structure is hypothesized. The negative sense single stranded RNA is of traditional 3’ to 5’ orientation. All the previously mentioned proteins are encoded by the RNA in the order of 3’-N-P-M-F-G-L- 5’12. Similar to all paramyxoviruses NiV RNA replication occurs in the cytoplasm. All but the P gene are monocystronic, in that they code for a single protein. The P gene also encoded for the C, V, and W proteins which play a role in the virulence of NiV. Interferons are released by host cells when under attack by a pathogen which enables intercellular communication. The intercellular communication is necessary for the triggering of immune cells which get rid of the pathogen. C, V, and W proteins, encoded by the P gene, have anti-interferon activity in that they block the transcription of interferon signalling.
_
Hendra virus (HeV) and Nipah virus (NiV) are reportedly the most deadly pathogens within the Paramyxoviridae family. In paramyxoviruses, viral attachment and membrane fusion are governed by the close interaction of the attachment proteins H/HN/G and the fusion protein F. The paramyxoviruses have two surface glycoproteins, the attachment (H/HN/G) and fusion (F) glycoproteins. These proteins work in concert; thus, in the case of NiV or HeV, the binding of G to a cellular receptor (ephrin B2/ephrin B3) induces a recently described conformational cascade in G that ultimately triggers F to execute pH-independent virus-cell or cell-cell membrane fusion. These two viruses bind the cellular entry receptors ephrin B2 and/or ephrin B3 via the viral attachment glycoprotein G, and the concerted efforts of G and the viral fusion glycoprotein F result in membrane fusion. Virus-cell and cell-cell membrane fusion are mediated by the viral fusion protein F, which for HeV and NiV requires the attachment protein G. Membrane fusion is essential for viral entry into host cells and for cell-cell fusion, a hallmark of the disease pathobiology. More detailed structural and functional analyses of the Hendra virus G glycoprotein may help us improve vaccine approaches and our understanding of HeV and NiV pathobiology.
_
The studies conducted by Tanimura et al. indicated that NiV is highly pathogenic to chicken embryos and that the chicken embryo represented a useful model for studying the vascular and neuronal tropisms of NiV. The virus also grows well in many of the mammalian cells but the rate of growth and patterns of cytopathic effect (CPE) produced in all culture varies with the type of mammalian cells used. With Vero cells, CPE could be visible by 5th–7th day post-inoculation of clinical samples; in subsequent passages, a complete CPE is observed by 24 h post-inoculation. It produces high viral titre (108 infectious particles per mL) in Vero cells at full CPE. The progression of the CPE also shows inclusions of viral nucleocapsids within the cytoplasm, budding of the nucleocapsid at the plasma membrane and pleomorphic extracellular enveloped virus particles filled with collection of tangled viral nucleocapsids.
_
Structure of the Nipah virus virion core:
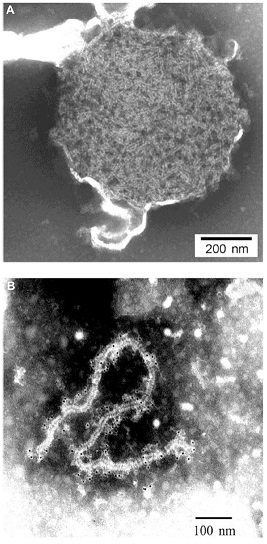
(A) Nucleocapsid core with residual envelope; (B) herringbone-shaped fragment of the nucleocapsid with immunogold labeling of P proteins. Nipah and Hendra viruses exhibit typical morphology of paramyxoviruses when examined by electron microscopy (EM), with a helical nucleocapsid structure surrounded by a membrane derived from the plasma membrane of the cell from which the viruses bud. In electron microscopic studies, negative stain preparations revealed nucleocapsids with the typical ‘herringbone’ appearance that is characteristic for paramyxoviruses produced by the association of the nucleocapsid protein with genomic RNA. Virus particles vary in size from 120 to 500 nm. Hendra virus (HeV) and Nipah virus (NiV) are enveloped, pleomorphic virions with diameters averaging about 500 nm, but may range from about 40 to 1900 nm in length. Thin-section EM studies of infected cells revealed filamentous nucleocapsids within cytoplasmic inclusions incorporated into virions budding from the plasma membrane. Pleomorphic extracellular virus particles, with an average diameter of 500 nm, and fine surface projections, were sporadically seen. In contrast to Nipah virus, which has only a single layer of surface projections, Hendra virus appears double-fringed, caused by projections on the surface of the viral envelope.
The nucleocapsid interacts with matrix (M) protein, located at the inner surface of the virion’s lipid envelope. The M protein, which drives the virus budding, also interacts with the cytoplasmic tale of the fusion (F) protein, thus stabilizing the virion. The F protein (type I membrane protein) and the attachment G glycoprotein (type II membrane protein) embedded in the lipid bilayer of the envelope form projections on the virion surface and elicit the development of neutralizing antibodies in all infected hosts, including bats. That makes the F and G proteins primary candidates as immunizing antigens in vaccine development. Infected hosts generally produce good non-neutralizing antibody titers against the N protein, making detection of anti-N antibodies suitable for enzyme-linked immunosorbent assay or Luminex assay, and of interest for the differentiating infected from vaccinated animals (DIVA) strategy in veterinary vaccine development. Location of the N gene at the very 3′ end of the genome makes it a sensitive target for RNA detection in infected cells due to the abundance of the transcripts from it.
_
Henipavirus fusion is a pH-independent process, but it requires proteolytic cleavage of the F protein (F0) into two subunits (F1 and F2). The F0 precursor expressed on the cell surface is endocytosed in clathrin-coated vesicles, cleaved by endosomal proteases known as cathepsins, and the functionally mature F protein is transported back to the plasma membrane, where it forms trimers. Direct plasma membrane fusion and macropinocytosis were identified as the henipavirus mode of cell entry, and this may be cell type specific. Plasma membrane fusion of infected cells with uninfected ones further facilitates virus spread throughout the host, and contributes to the pathology of henipavirus disease. Epithelial and endothelial syncytia, and multinucleated cells (mostly macrophages and dendritic cells), were observed in the tissues of humans and other host species.
Henipavirus fusion by the F protein also requires, in addition to cleavage of the F protein, activation of this protein by the second viral glycoprotein, the G protein. Henipavirus G glycoproteins form covalently linked dimers noncovalently associated into tetramers. In contrast to attachment glycoproteins of other paramyxoviruses, henipavirus G proteins lack both hemagglutinin and neuraminidase activities. HeV and NiV glycoprotein G is the virus attachment protein recognizing ephrin B2 and ephrin B3 as cellular receptors, and it may also bind with lower affinity to a C-type lectin on endothelial cells in the lymph nodes and liver. Ephrin B2 is important in mammalian host development and it is a highly conserved protein across mammalian species, expressed on lymphocytes, neurons, smooth muscle cells, and endothelial cells surrounding small arteries, corresponding with virus distribution in an infected host as determined by immunohistochemistry. The cellular function of ephrin B2 is to regulate processes such as neurogenesis, angiogenesis, proliferation, and remodeling, as well as immune activation and bone formation. HeV G protein has a somewhat lower affinity for ephrin B3 compared to the NiV G protein. This may play a role in the somewhat different pathology in human infections, where NiV with its higher affinity for this receptor than HeV, causes fatal neurological disease with severe brain stem dysfunction, although the last human fatal case of HeV had extensive brain involvement. Interestingly, in experimentally infected swine, HeV invasion of the central nervous system (CNS) was limited in the early stages to the olfactory bulb alone, and with a lower inoculation dose, HeV did not reach the CNS (Pickering B, Weingartl HM, 2015). Using ubiquitous and conserved proteins as receptors, henipaviruses not only have a wide host range, but they also infect a wide range of tissues and organs within each individual host species.
The P gene of henipaviruses encodes three nonstructural proteins (C, V, and W) in addition to the phosphoprotein P. The P protein is critical for virus replication, and through its interaction with cellular proteins, it also modulates cell signaling. Some of the functions encoded in the N terminus of the protein are shared with the V and W proteins due to an identical N-terminal portion of the three proteins. The V protein is produced by the cotranscriptional addition of one nontemplated G at the editing site, and the W protein by insertion of two nontemplated Gs. The proteins are located in the cytoplasm, except for the W protein being detected also in the nucleus in some cell types. The C protein is encoded by a separate internal open reading frame within the P gene, and it localizes predominantly into the perinuclear region. The P, W, and V proteins hinder the interferon JAK–STAT signaling pathway by binding to STAT1 and preventing its translocation into the nucleus. Nuclear localization of NiV W impairs the TLR3/TRIF pathway by blocking TRIF-mediated activation of interferon regulatory factor (IRF)-3 responsive promoter, ultimately interfering with the induction of interferon (IFN)-β and other molecules controlled by this pathway. NiV V proteins bind to the MDA5 helicase along with LGP2 to suppress RIG-I-like (RLR) signaling, thereby inhibiting the downstream signaling events also leading to IFN-β synthesis. NiV C exhibits inhibitory activity against TRL7/9-dependent IFN-α induction by binding to IKKs and inhibiting phosphorylation of IRF-7,102 as well as influencing IFN-β and antiviral gene expression. A study by Mathieu et al suggested that the C protein can regulate cytokine balance in transfected cells. The nonstructural proteins are involved in the NiV life cycle by regulating replication and evading the innate immune response, as confirmed by in vivo reverse genetics studies in animal models. The evasion of the IFN system appears to be cell specific, since NiV will induce IFN type I and other innate cytokines in endothelial cells, an important in vivo target for henipaviruses, and there is the possibility that cells may employ alternative pathways to establish an antiviral state. The molecular aspects of henipavirus replication and its interaction with the host cell are – with some differences between HeV and NiV, and between individual nonreservoir host species – reflected in the clinical disease, pathogenesis, pathology, and immune response.
_
Reproduction of nipah virus:
It can be assumed, because NiV is a paramyxovirus it acts very similar to other viruses within the family. The reproduction mechanism is very similar to influenza. The virion binds and fuses to the surface of a host cell via the F and G proteins. The lipid bi-layers are then melted and the viral nucleocapsid is released into the host cell. The negative sense viral RNA is transcribed to mRNA which acts as a template for more negative sense viral RNA. The viral RNA is used to make the necessary proteins (N, P, M, F, G, L, C, and V, W) which congregate near the cell membrane. Once all the necessary proteins are assembled a new viral cell will bud off and infect other host cells. The new viral cells are able to fuse together and create a huge multinucleated cell called syncytia. A major different between the reproduction of paramyxoviruses and influenza is paramyxovirues are strictly reproduced in the cytoplasm.
_
Cultured Nipah virus:
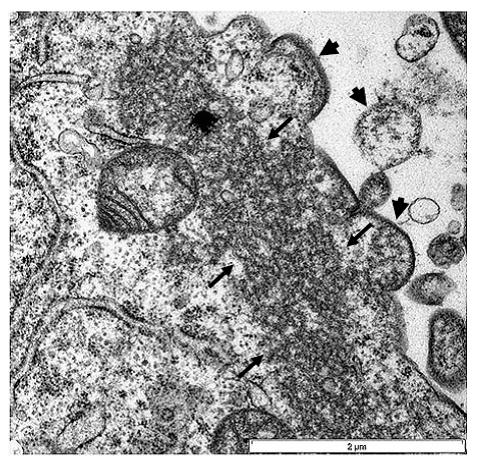
The small arrows indicate the nucleocapsids and the large arrow heads indicate budding virions developing from the cell. Picture scale is 2um. HeV and NiV particles are enveloped and pleomorphic with spherical or filamentous forms observed by electron microscopy.
_
Mutation rate of NiV:
NiV is an RNA virus and a zoonotic virus: RNA viruses have a high mutation rate which enables them to keep a leg up on both vaccines and host immune systems. Zoonotic viruses also have a high mutation rate. Because NiV is both of these, it is hypothesized it has an extremely high rate of mutation. Mutation rate is typically defined as the average number of errors created in genomes of viral progeny, per base, per replication cycle (mut/nuc/rep). Viruses possess mutation rates that are orders of magnitude higher than any other replicating entity. These rates range from approximately 1.5 × 10−3 mut/nuc/rep in the RNA bacteriophage Qβ (Batschelet et al., 1976) to ∼10−8 mut/nuc/rep in the DNA virus Herpes simplex (Drake and Hwang, 2005). While mutations create raw genetic diversity, it is the coupled action of mutation and selection that will determine which mutations will persist in the viral population. The rate at which mutations fix in a population is termed the substitution rate, or evolutionary rate, which is measured by comparing the genomes of different isolates of a virus collected at several different time points (Duffy et al., 2008). RNA viruses possess much higher substitution rates than DNA viruses, ranging from 0.01 substitutions per site per year (sub/site/yr) in the RNA poliovirus type 1 to 7 × 10−7 in the DNA virus monkeypox. As suggested by theory, in most viruses substitution rates correlate well with mutation rates. This suggests that the short-term mutation rate is an important determinant of the rate of long-term molecular evolution. The mean value of Nipah virus N gene evolutionary rate is 6.5 × 10−4 substitution per site per year. Influenza A virus evolutionary rate ranged from a minimum of 1.43 × 10−3 substitution per site per year (H3N8) to a maximum of 11.62 × 10−3 (H7N7).
__
Resistance to physical & chemical action, and survival of nipah virus:
As other animal Paramyxovirus, the virus is inactivated by 60°C for 60 minutes. However the virus was not completely eliminated in artificial palm sap held at 70°C for 1 hour. It is stable between pH 4.0 and 10.0. It survives for long periods in favourable conditions, for hours to days in fruit bat urine, contaminated fruit juice and date palm sap. It is susceptible to common soaps and disinfectants. Lipid solvents, such as alcohol and ether, and sodium hypochlorite solutions were used effectively in outbreaks for disinfection.
_____
HeV and NiV viruses, entry and tropism:
Genomic sequence analysis of many HeV isolates obtained from horses, a human case, and pteropid bats, have shown them to be ∼99% identical. In the initial Malaysian outbreak of NiV, both pig and human isolates were genetically highly similar to those obtained years later from bats. However, some diversity among NiV isolates can be noted when comparing Malaysian isolates to NiV isolates from other areas of Southeast Asia, with NiV-Bangladesh sharing ∼92% identity with NiV-Malaysia, and a third lineage of NiV was isolated from Lyle’s flying fox (P. lylei) in Cambodia that is more closely related to NiV-Malaysia than to NiV-Bangladesh.
HeV and NiV are also distinguished by their exceptionally broad species tropism and in addition to pteropid bats, natural or experimental infection has been documented in pigs, horses, cats, dogs, guinea pigs, mice, hamsters, ferrets, squirrel monkeys and African green monkeys and humans. NiV can also productively infect chicken embryos spanning 7 orders (6 mammalian and one avian).
The major determinant of species and cellular tropism of HeV and NiV is derived from the functions of the viral envelope glycoproteins (G and F) which are the mediators of virus attachment and host cell infection. The HeV and NiV G glycoprotein bind to the host cell membrane proteins ephrin-B2 and ephrin-B3. The ephrin-B2 and -B3 molecules are members of a large family of cell surface expressed glycoprotein ligands that bind to Eph receptors and are highly sequence conserved across known susceptible hosts with amino acid identities ranging from 95-98%. Ephrin-B2 expression is prominent in arteries, arterioles and capillaries in multiple organs and tissues while ephrin-B3 is found predominantly in the nervous system and the vasculature. Their identification as major receptors for HeV and NiV has helped clarify their broad species and tissue tropisms and pathogenic features observed in both animals and humans.
_____
_____
Phylogenetic and evolutionary analysis represents promising tools to evidence NiV epidemics, to study their origin and evolution and finally to act with effective preventive measure:
Nipah virus (NiV) is a member of the genus Henipavirus of the family Paramyxoviridae, characterized by high pathogenicity and endemic in South Asia. It is classified as a Biosafety Level-4 (BSL-4) agent. The case-fatality varies from 40% to 100% depending on the severity of the disease and on the availability of adequate healthcare facilities. At present no antiviral drugs are available for NiV disease and the treatment is just supportive. NiV infection can be considered an emerging disease and a public health problem as a consequence of the lack of effective vaccines and therapies and of the evidence that NiV can infect pigs. Phylogenetic and evolutionary analyses can represent very useful tools to elucidate the epidemiology and the temporal origin of this virus. Moreover, these analyses, especially the evolutionary analysis, could be advantageous to develop new therapy, vaccine and prevention strategies. The circulation of NiV may be influenced by the presence of genetic polymorphisms along the virus genome. As a consequence, the antigenic variability is possible and may play an important role in the ability of the virus to escape the host immune response. Recently, it was reported that monocytes, natural killer cells and CD4+/CD8+ T cells support Nipah virus replication, which may facilitate dissemination of the virus during pathogenesis.
________
Viral interference may be a useful mechanism to combat viral infections, a 2007 study:
As viral interference may be a useful mechanism to combat viral infections, possibly including those with NiV and HeV, authors generated cell lines expressing the viral proteins G and F. After exposure to henipaviruses, transgenic cells expressing the NiV G attachment glycoprotein were resistant to infection with NiV and HeV. The results demonstrated that a viral interference system can be established for these members of the family Paramyxoviridae and that expression of the attachment glycoprotein did not result in cell-surface downregulation of the viral receptors.
_____
The C, V and W proteins of Nipah virus inhibit minigenome replication, a 2008 study:
As very little is known about the replication of the henipaviruses, the goal of this research was to investigate the roles that the C, V and W proteins of NiV play in viral transcription and replication. Here, authors demonstrate that the C, V and W proteins of NiV inhibit NiV minigenome replication. The NiV C, V and W proteins also inhibited replication of a measles virus (MV) minigenome, and the C and V proteins of MV inhibited NiV minigenome replication. However, the accessory proteins of NiV and MV were not able to inhibit human parainfluenzavirus 3 (hPIV3) minigenome replication. Conversely, the C protein of hPIV3 inhibited minigenome replication of both NiV and MV. These findings suggest that these divergent viral proteins share common functional properties, perhaps through interaction with a common host cell protein(s).
_____
_____
Epidemiology of Nipah virus:
The epidemiology of Nipah and Hendra viruses has not been fully elucidated. A similarity exists between the epidemiology of each of these two viruses, such as the Pteropus fruit bat as the natural host for both viruses. For both Hendra and Nipah viruses, it is presumed that horses and pigs that have acted as an intermediary host to humans had been infected by indirect contact with pteropid bats endemic in these regions, although this has not been experimentally proven. However, there are also differences in their epidemiology. Hendra virus was first described along the coastal regions of Australia, and to date has caused illnesses in 5 humans with 2 deaths, in infections that were acquired by close contact with ill horses infected with the virus. Nipah virus infections in humans have been described in Malaysia, Singapore, Bangladesh, and India, and have been identified in bats in Cambodia and Thailand. Until 2004, cases occurred only in clusters, but sporadic cases have been identified more recently through active surveillance (Anon., 2004). Overall, Nipah virus has caused at least 365 human deaths to date. Direct contact with infected pigs was primarily responsible for the outbreak in Malaysia, although in Bangladesh the epidemiology was less well-defined, with some evidence for person-to-person transmission.
_
The outbreak in Malaysia was initially thought to be Japanese encephalitis (JE), which is endemic in Asia. However, certain features were not consistent with this diagnosis: the occurrence of encephalitis among adults rather than children, the clustering of cases in the same household, and a history of illness in pigs belonging to the affected farmers. Furthermore, a high number of patients had been vaccinated against JE. The growth of large intensively managed commercial pig farms in Malaysia with fruit trees on the farm created an environment where bats could drop partially eaten fruit contaminated with NiV laden bat saliva into pig stalls. The pigs could eat the fruit, become infected with NiV, and efficiently transmit virus to other pigs because of the dense pig population on the farms, frequent respiratory shedding of the virus among infected pigs (Middleton et al., 2002), and the pigs’ high birth rate that regularly brought newly susceptible young pigs into the population at risk (Epstein et al., 2006). As bats fed and roosted in the trees, nearby livestock became infected with Nipah virus, which eventually spread to farm labourers. It is assumed that the geographic distribution of Henipaviruses overlaps with that of Pteropus category. This hypothesis was reinforced with the evidence of Henipavirus infection in Pteropus bats from Australia, Bangladesh, Cambodia, China, India, Indonesia, Madagascar, Malaysia, Papua New Guinea, Thailand and Timor-Leste. Furthermore, the detection of antibodies against Nipah and Hendra viruses in strawcoloured fruit bat (Eidolon helvum), indicates that these viruses might be present within the geographic distribution of Pteropodidae bats, not only in Asia, but extended to Africa, Arabian peninsula coast, Middle-East, Cyprus and Southern Turkey.
_
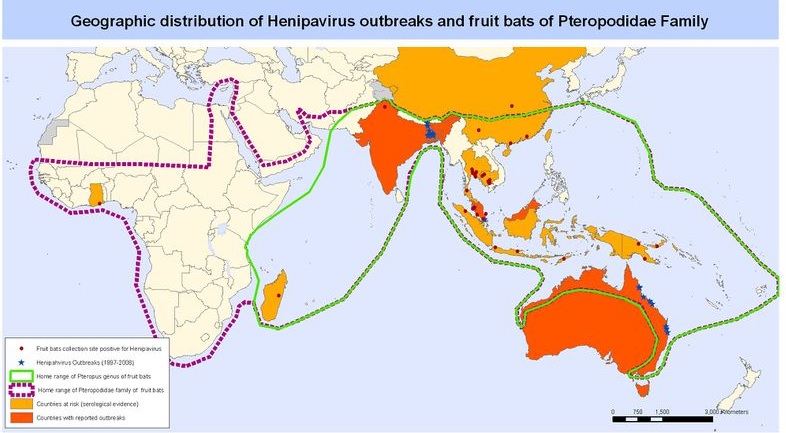
This is a map of Southeast Asia and Africa and the distribution of Henipavirus outbreaks and Pteropus species. Red circles indicate Pteropus collection sites who found bats positively infected with NiV, blue stars indicate Henipavirus outbreaks, green line indicates the entire range of the Pteropus genus, purple dashed line indicated the entire range of pteropodidae family, yellow countries are at high risk for outbreaks, and orange countries have reported outbreaks. (WHO 2001)
_
Chronology of Nipah Outbreaks:
Although Nipah virus has caused relatively few outbreaks, it infects a wide range of animals and causes severe disease and death in people, making it a public health concern. Nipah virus was first recognized in 1998 during an outbreak among pig farmers in Malaysia. Since then, there have been various outbreaks, all in South Asia. The chronology of outbreaks due to Nipah virus is summarized in table below.
Chronology of outbreaks due to Nipah virus (1998-2018):
| Year | Country | State or District | Cases | Deaths | Case fatality |
| 1998-1999 | Malaysia | Perak, Selangor, Negeri Sembilan states | 265 | 105 | 40% |
| 1999 | Singapore | Singapore | 11 | 1 | 9% |
| 2001 | India | Siliguri district, West Bengal | 66 | 49 | 74% |
| 2001 | Bangladesh | Meherpur district | 13 | 9 | 69% |
| 2003 | Bangladesh | Naogaon district | 12 | 8 | 67% |
| 2004 | Bangladesh | Faridpur and Rajbari districts | 67 | 50 | 75% |
| 2005 | Bangladesh | Tangail dstrict | 12 | 11 | 92% |
| 2007 | Bangladesh | Thakurgaon, Naoga and Kushtia districts | 18 | 9 | 50% |
| 2007 | India | Nadia district, West Bengal | 5 | 5 | 100% |
| 2008 | Bangladesh | Manikgonj, Rajbari and Faridpur district | 11 | 9 | 82% |
| 2009 | Bangladesh | Rajbari, Gaibandha, Rangpur and Nilphamari districts | 4 | 1 | 25% |
| 2010 | Bangladesh | Faridpur, Rajbari, Gopalganj and Madaripur districts | 16 | 14 | 88% |
| 2011 | Bangladesh | Lalmonirhat, Dinajpur, Comilla, Nilphamari and Rangpur districts | 44 | 40 | 91% |
| 2012 | Bangladesh | Joypurhat Rajshahi, Natore, Rajbari and Gopalganj districts | 12 | 10 | 83% |
| 2013 | Bangladesh | Gaibandha, Jhinaidaha, Kurigram, Kushtia, Magura, Manikgonj, Mymenshingh, Naogaon, Natore, Nilphamari, Pabna, Rajbari and Rajshahi districts | 24 | 21 | 87% |
| 2015 | Bangladesh | Nilphamari, Ponchoghor, Faridpur, Magura, Naugaon and Rajbari districts | 9 | 6 | 67% |
| 2018 | India | Kozhikode and neighboring Malappuram district, Kerala | 19 | 17 | 89% |
Note: Table above depicts not only chronology of outbreaks but also mortality rate.
_
The initial outbreak of encephalitis, later discovered to be a result of NiV, and first epicenter occurred in September of 1998 in several pig farms in a small town called Ipoh in northern Malaysia. NiV is thought to have been transmitted through southern Malaysia by the transference of infected pigs to a second epicenter located around Kampung Sungai Nipah. This outbreak resulted in 265 infected patients with 105 deaths, a fatality rate of 40%. The main transmission of this outbreak is thought to be from the reservoir host to an intermediate host, to humans. This outbreak also resulted in a huge death rate of the pig population in Malaysia and Singapore. Since the outbreak in 1998, a dozen outbreaks have since occurred in Bangladesh and India starting in 2001. These outbreaks have resulted in more respiratory disease and a fatality rate of up to 92% which have lead scientists to suspect a different strand of Nipah virus as the culprit. The transmission is thought to be directly from the reservoir host to humans, along with a higher human to human transmission rate. Overall in Bangladesh, there had been 188 infections with 146 deaths with a fatality rate of 77%. The most recent outbreak occurred in India in May 2018 resulting in 19 infections and 17 deaths. Clinical material obtained during the Siliguri outbreak was retrospectively analyzed for evidence of NiV infection. Nipah virus-specific immunoglobulin M (IgM) and IgG antibodies were detected in 9 out of 18 patients. Reverse transcription-polymerase chain reaction (RT-PCR) assays detected RNA from NiV in urine samples from 5 patients. A second outbreak was reported in 2007 in Nadia district of West Bengal. Thirty cases of fever with acute respiratory distress and/or neurological symptoms were reported and five cases were fatal. All five fatal cases were found to be positive for NiV by RT-PCR.
_
In contrast to the Malaysia-Singapore outbreak, which could be coherently explained by a single or perhaps a few transmissions of NiV from an infected bat to pigs, leading to a porcine epidemic which in turn led to a human epidemic (Epstein et al., 2006); in Bangladesh NiV transmission from bats to human is repeated and ongoing. The diversity of NiV strains recovered from Bangladesh also supports multiple introductions of the virus from bats into human populations even within a single year. Among 4 NiV isolates from human NiV cases in 2004, the sequences of the nucleoprotein open reading frames of the isolates differed by 0.9% in nucleotide homology, in contrast to the sequences obtained from all of the human cases in Malaysia which were nearly identical to each other (AbuBakar et al., 2004; Chan et al., 2001; Harcourt et al., 2005). The clinical presentation of NiV infection in Bangladesh differed from Malaysia. In Bangladesh, severe respiratory disease is more common, with 62% of cases having cough, 69% developing respiratory difficulty, and available chest radiographs showing diffuse bilateral opacities covering the majority of the lung fields (Hossain et al., 2008). By contrast, in Malaysia, 14% of patients had a non-productive cough on presentation; only 6% of chest radiographs were abnormal and these abnormalities were mild and focal (Goh et al., 2000). The case fatality rate was higher in Bangladesh at 73%, compared with 40% from Malaysia (Goh et al., 2000; Hossain et al., 2008), but much of this difference results from the more sophisticated clinical care provided in Malaysia. One-half of Malaysian Nipah patients received mechanical ventilatory support compared to a single patient (1%) in Bangladesh (Goh et al., 2000).
_
Chronology of nipah and hendra virus is depicted in the figure below:
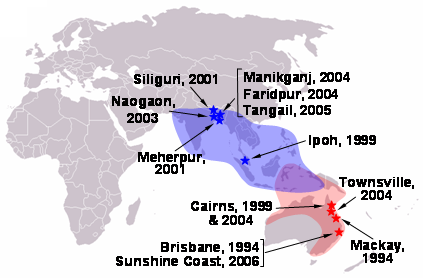
_
The Nipah virus infection has become endemic in Bangladesh, causing regularly outbreaks, in particular in districts where date palm sap is seasonally produced. Various reports referred to outbreaks since 2001. The NiV epidemiology in Bangladesh can be schematically referred to the so called “Nipah belt” (figure below), corresponding to northern-central districts of the country where date palm sap collection is also common. Nipah virus outbreaks in Bangladesh are most common during the winter season.
Nipah belt in Bangladesh:
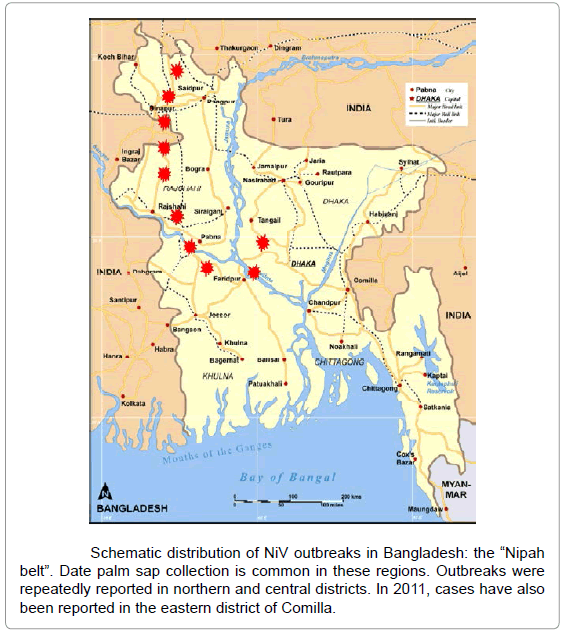
_
Nipah virus outbreaks have been reported in Malaysia, Singapore, Bangladesh and India. The highest mortality due to Nipah virus infection has occurred in Bangladesh. In Bangladesh, the outbreaks are typically seen in winter season. Nipah virus first appeared in Malaysia in 1998 in peninsular Malaysia in pigs and pig farmers. By mid-1999, more than 265 human cases of encephalitis, including 105 deaths, had been reported in Malaysia, and 11 cases of either encephalitis or respiratory illness with one fatality were reported in Singapore. Infected pigs from Malaysia were transported to Singapore resulting in outbreak in Singapore. In 2001, Nipah virus was reported from Meherpur District, Bangladesh and Siliguri, India. The outbreak again appeared in 2003, 2004 and 2005 in Naogaon District, Manikganj District, Rajbari District, Faridpur District and Tangail District. In Bangladesh, there were also outbreaks in subsequent years. In May 2018, an outbreak was reported in the Kozhikode and Malappuram districts of Kerala, India, eighteen deaths have been recorded, including one healthcare worker. Those who have died are mainly from the districts of Kozhikode and Malappuram, including a 31-year-old nurse, who was treating patients infected with the virus.
_
Species Affected:
Fruit bats of the genus Pteropus (flying foxes) are the main reservoir hosts for Nipah virus. P. vampyrus, the Malayan flying fox, and P. hypomelanus, the island flying fox, are known to carry this virus in Malaysia. P. giganteus is thought to be an important host in Bangladesh and India and possibly other locations. Although live virus has not yet been isolated from this species, Nipah virus RNA has been detected and many bats are seropositive. Nipah virus also occurs in P. lylei in Thailand and Cambodia, and P. poliocephalus has been infected experimentally. Viral RNA and/or antibodies have been found in a few other species of fruit or insectivorous bats, although their significance is unclear. Many domesticated mammals seem to be susceptible to Nipah virus. This virus can be maintained in pig populations, but other domesticated animals appear to be incidental (spill-over) hosts. Sick goats, dogs, cats and horses were observed in the outbreak area in Malaysia, and infections in dogs, a cat, a horse and goats were confirmed by immunohistochemistry. Sheep might also have been affected, but there are no confirmatory data, and no evidence of infections could be found in rats. Nipah virus seems to have affected horses in the Philippines in 2014, based on clinical signs in the horses and epidemiological links to human patients; however, no tissues were available from the horses for confirmation. Several cats and a dog that had eaten tissues from sick horses in the Philippines also died, and seropositive dogs were reported in the outbreak area. Another study reported seropositive cattle, pigs and goats in Bangladesh; however, these antibodies did not neutralize Nipah virus, and could have been caused by related henipaviruses. Experimental infections with Nipah virus have established in pigs, cats, ferrets, nonhuman primates, guinea pigs, golden hamsters (Mesocricetus auratus) and mice.
Species Susceptible to NiV:
Humans, pigs, bats, dogs, cats, goats and horses are susceptible to NiV infection. NiV infection has been reported also in sheep, but the observation could not be further confirmed and remains controversial. Clinical disease can be observed in experimental conditions in ferret (Mustela putorius furo), guinea pig (Cavia porcellus), squirrel monkey (Saimiri sciureus), African green monkey (Chlorocebus aethiops) , hamster (Cricetinae), and in suckling mouse (Mus musculus).
_____
_____
Natural host and reservoir of NiV:
- Primary reservoir for Nipah virus is fruit bats of the genus Pteropus
- Domestic swine are extremely susceptible to infection; act as amplifying host
- Infections have also been reported in dogs, cats, horses and goats
_
Natural Host:
Fruit bats (Macrochiroptera) of the family Pteropodidae- particularly species belonging to the Pteropus genus–are the natural hosts for Nipah virus. There is no apparent disease in fruit bats. Bats belonging to the genus Pteropus are widely distributed. They live in the tropics and subtropics of Asia, including the Indian subcontinent, Australia, Indonesia, Madagascar, and a number of remote oceanic islands in both the Indian and Pacific Oceans. A major breakthrough was achieved by Dr Lam and his team who studied a large bat colony on Tioman island, off the eastern coast of West Malaysia. After collecting over 1,000 bat urine samples, the virus was detected in the urine of a species of flying fox. The virus was also found in a piece of fruit that had been partially eaten by a bat. This implied the presence of the virus in the animal’s saliva. Because many Malaysian pig farms have fruit trees, it is easy enough to imagine how the virus can be transmitted from bats. Pigs probably serve as amplifying hosts after ingesting infected bat urine, saliva, or discarded food items. Dr Lam suggested that pig farmers stop growing or maintaining fruit trees in order to prevent a future outbreak.
_
Fruit bat:

Among the genus Pteropus, the Indian Flying Fox (Pteropus giganteus) (wingspan 1.5 m and up to 1.2 kg) and the relatively smaller Greater short-nosed fruit bat or Short-nosed Indian fruit bat (Cynopterus sphinx) (wingspan 48 cm), widespread and very common species in South Asia, have been identified as the main natural reservoir. Various other pteroid bats have been recognized NiV host carriers. The grey-headed flying fox (Pteropus poliocephalus) and the black flying-fox (Pteropus alecto), both Pteropus spp. occurring in Malaysia were found seropositive for NiV. Neutralizing antibodies, and the virus has been isolated from the small flying fox or variable flying fox (Pteropus hypomelanus) and the large flying fox (Pteropus vampyrus). NiV has been isolated from urine of Lyle’s flying fox (Pteropus lylei) in Cambodia. The Pteropus conspicillatus, or spectacled flying fox, lives in Southeast Asia and Northern Australia. This species is also a reservoir host of NiV.
_
Yob et al. and Chua have confirmed that fruit bats (popularly known as flying foxes) found in Malaysia, P. vampyrus and P. hypomelanus, are the natural reservoir hosts of NiV. The virus is assumed to have been introduced into the swine population from bat species of Pteropus. Malaysia has diverse bat fauna, with at least13 species of fruit bats, including two species of flying foxes (Pteropid bats) and more than 60 species of insectivorous bats. A serological survey during the 1999 outbreak demonstrated neutralizing antibodies in 5 species of bats, 4 species of fruit bats and one species of insectivorous bats. This included 31 % positive antibody status to Pteropus hypomelanus (island flying foxes) and 17 % positive antibody status to Pteropus vampyrus (Malayan flying foxes) in Peninsular Malaysia . Serologic and RT-PCR testing of blood and urine from Pteropid fruit bats in Malaysia and Cambodia showed NiV infection, which suggested that these animals were reservoir hosts. The geographic range of Pteropus includes all of southeast and south Asia and several other species have been found with antibodies to NiV. The virus has been found in excretions of Pteropid bats, Chua et al. isolated NiV from urine of bats in Malaysia.
_
Fruit bat range:
Figure above shows range of Pteropus bats.
Flying foxes occur across South-east Asia. The world distribution of flying foxes (genus Pteropus) extends from the west Indian Ocean islands of Mauritius, Madagascar and Comoro, along the sub-Himalayan region of Pakistan and India, through South-east Asia, the Philippines, Indonesia, New Guinea, the South-west Pacific Islands (to the Cook Islands), and Australia (Figure below).
_
Map showing locations of outbreaks of Nipah and Hendra virus as well as the range of Pteropus bats as of 2014.
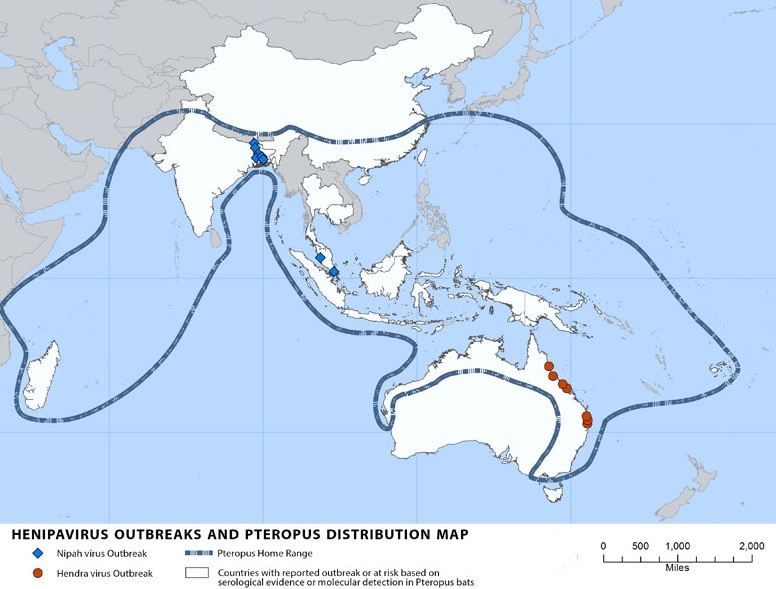
_
There are about 60 species of flying foxes in total. Flying foxes range in body weight from 300 g to over 1 kg, and in wingspan from 600 mm to 1.7 m. They are the largest bats in the world, do not echolocate but navigate at night by eyesight and their keen sense of smell. Females usually have only one young a year, after a six-month pregnancy. The young are independent after about three months. All flying fox species eat fruits, flowers or pollen, and roost communally in trees (Hall & Richards 2000). Flying foxes are nomadic, known to travel over considerable distances. Radiotracking studies in eastern Australia have shown individuals to undertake periodic movements of up to 600 km (Eby 1991). Where the distributions of different species overlap, roosts are shared. Thus the potential exists for interaction between flying fox populations across the region.
_
Suspicion of bats as natural hosts for the zoonotic paramyxoviruses began after neutralizing antibodies to Hendra virus were found in 4 species of pteropid bats in Australia (Young et al., 1996). Hendra virus has since been isolated from reproductive tissue from P. poliocephalus and P. alecto (Halpin et al., 2000). Hendra virus has been demonstrated in Australian bats from the northern city of Darwin down to Melbourne as well as in Papua New Guinea (Paterson et al., 1998). Extensive animal surveillance among other animals has not shown evidence of natural Hendra virus infection among horses or farm animals, or among more than 40 species of wildlife tested from Queensland (Rogers et al., 1996; Ward et al., 1996). Antibodies to Nipah virus have been found in 9–25% of pteropid bats in Malaysia, Cambodia, Thailand, and Bangladesh, (Yob et al., 2001; Hsu et al., 2004; Reynes et al., 2005; Wacharapluesadee et al., 2005). Neutralizing antibodies to Nipah virus were found in P. hypomelanus and P. vampyrus, the two pteropid species in Peninsular Malaysia; in Cambodia, antibodies were found in a third species, P. lylei; while in Thailand, antibodies were present in all three species. In Bangladesh, P. giagnteus bats, a more common species in that region, were found to have neutralizing antibodies to Nipah virus. Neutralizing antibodies to Nipah virus have also been found in other frugivorous and insectivorous bat genera including Eonycteris, Cynopterus, Scotophilus, and Hipposideros, although in a lower proportion than in Pteropus spp. (Yob et al., 2001;Wacharapluesadee et al., 2005; ); whether these bats are also considered natural hosts and the significance of these findings have yet to be determined. Isolation of Nipah virus from the bats has proven to be difficult, but the virus was isolated from 3 of 263 pooled bat urine samples in Malaysia, and 2 of 769 urine samples in Cambodia (Chua et al., 2002; Reynes et al., 2005).
_
Large fruit bats of the genus Pteropus appear to be the natural reservoir of NiV. In Malaysia the seroprevalence of neutralizing antibodies to NiV in colonies of Pteropus vampyrus and Pteropus hypomelanus ranged from 7% to 58% (Yob et al., 2001; Daszak et al., 2006). Antibodies against henipaviruses have been identified in Pteropus bats wherever they have been tested including Cambodia, Thailand, India, Bangladesh, and Madagascar (Epstein et al., 2004; Hsu et al., 2004; Iehle et al., 2007; Reynes et al., 2005; Wacharapluesadee et al., 2005). NiV was isolated from urine specimens collected underneath a P. hypomelanus roost and from partially eaten fruit dropped during feeding activity in Malaysia (Chua et al., 2002), from urine collected underneath a Pteropus lylei roost in Cambodia (Reynes et al., 2005), and from saliva and urine of P. lylei in Thailand (Wacharapluesadee, 2005). In the 2007 Bangladesh outbreak, 50% of Pteropus bats sampled in the region 1 month after the outbreak had antibodies to Nipah virus, suggesting bats as a possible zoonotic source. Experimental infection of Pteropus bats with NiV does not cause illness in the bats (Middleton et al., 2007). Surveys of rodents and other animals have not identified other wildlife reservoirs for NiV (Hsu et al., 2004; Yob et al., 2001). Over 50 species of Pteropus bats live in South and South East Asia (Nowak, 1994). Pteropus giganteus, the only Pteropus species found in Bangladesh, is widely distributed across the country and frequently has antibody to NiV (Bates and Harrison, 1997; Hsu et al., 2004).
_
It has been confirmed that fruit bats were the source of the deadly nipah virus that killed 17 people in the Kerala in May 2018. Tests conducted by Indian Council of Medical Research (ICMR) confirmed fruit bats from Changaroth panchayat in Kerala’s Kozhikode district, as the source of the outbreak. The 21 bats trapped in the first round and tested were insectivores that did not carry the Nipah Virus; out of the 51 Pteropus giganteus bats sampled by the NIV team from the Kozhikode epicentre of the outbreak, 11 had genetic material from the Nipah virus. These portions of the viral genome were detected through a test called Reverse Transcriptase Polymerase Chain Reaction (RT-PCR), and were 99.68% similar to the virus in Kerala’s patients. This indicates strongly that the bats were the carriers in the Kerala outbreak. The genome of the virus isolated from Kerala’s patients was similar to the Bangladeshi Nipah strain. However, this doesn’t mean that Bangladeshi travellers or bats brought the virus to Kerala.
_
The absence of evidence for Nipah in fruit bats does not rule out Nipah infection in fruit bats:
- It is difficult to find a bat that is infected. In a population of 100,000 bats only four or five could be carrying the Nipah virus.
- Bats may only be infectious for a week or two, and then they clear the virus and they are no longer infectious. So even if a bat has been infected by virus earlier, their samples may test negative later. The time window in which the bats are capable of passing on the infection to other animals or humans is quite small. In fact, the virus cannot be found in experimentally infected bats after a few weeks.
- The few bats in an infected population that could be shedding the virus may be doing so in low quantities and for a short duration. And the shedding of the virus in secretion is not uniform.
- In India, fruit bats have been tested for the Nipah virus. A 2012 study by researchers from the National Institute of Virology, Pune, and the US Centres for Disease Control and Prevention had found the virus in only one of 140 bats tested in Maharashtra and West Bengal. The other 139 bats were not found to have even the antibodies necessary to fight the virus. The paper noted the need for a systematic survey to understand the Nipah virus distribution. A 2008 paper had found that 20 of 41 bats tested in Haryana had antibodies to the Nipah virus. Apart from the difference in location, this disparity in the proportion of bats with antibodies to the virus is not surprising. So there are different proportions of bats in any given colony exposed to the Nipah virus. This proportion is always changing, and what such surveys reveal will only be a snapshot.
- Although there are hundreds of bats species, the research primarily implicates giant fruit bats, which travel quite a long distance to feed at night, up to fifty kilometers—thirty-one miles—from their roost. This means you don’t need to find giant fruit bats roosting where people were initially infected; they could have flown in from ten kilometers—six miles—away to eat some fruit or drink sap in that backyard the night someone was infected. That’s why adequate wildlife testing requires an understanding of bat ecology and behavior; it’s not simply a matter of grabbing the closest bats to the house of the infected person.
__
Table below shows distribution of bat species shown to have Nipah virus:
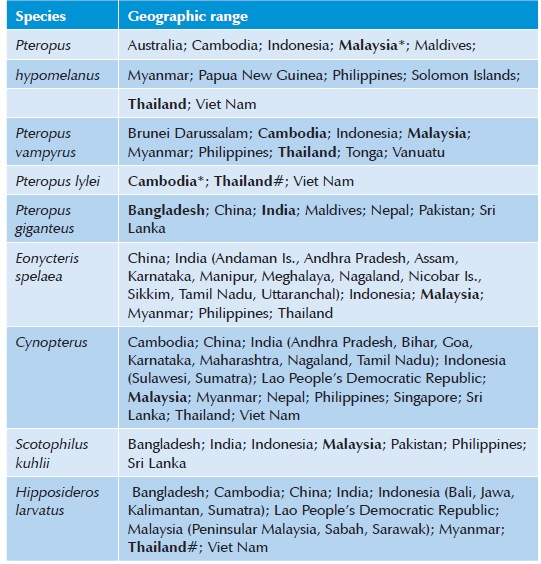
Bold, countries where Nipah virus infection in bats was demonstrated by antibody detection method.
*, countries where Nipah virus infection in bats was confirmed by isolation.
#, countries where Nipah virus infection in bats was confirmed by RNA detection.
_
Rahman et al. in 2010, reported the results of a prospective cohort study focused on a group of P. vampyrus flying foxes captured in two different locations in Malaysia. Authors showed that NiV detected in P. vampyrus differs from all known isolates from Malaysia for the amino acid changes at 44 positions. The phylogenetic analyses unequivocally showed that NiV P. vampyrus forms a monophyletic clade with other NiV isolates from Malaysia, but it differs from human, pig, and Pteropus hypomelanus bat isolates. When 56 NiV sequences from Pteropus lylei bats isolated in Thailand were included, NiV P. vampyrus phylogenetically grouped most closely with NiV P. lylei, and the monophyly of NiV sequences from Malaysia was lost. This close homology suggested that NiV is naturally transmitted between these two species. From this study, the presence of NiV diversity in isolates from P. lylei bats has also emerged. This diversity demonstrated that multiple strains co-circulate within populations and that the ecology and sympatry of Pteropus spp., not coevolutionary patterns, are determinant for the NiV strain diversity observed in reservoir hosts.
_
Serological evidences indicate that circulation of Henipaviruses in bats is not limited to species belonging to the genus Pteropus, but also extended to a wider range of both frugivorous and insectivorous bats. An example is represented by the Lesser Asiatic yellow house bat (Scotophilus kuhlii) (wingspan up to 5.2 cm, weight up to 22 gr), insectivorous bat (Microchiroptera) of the genus Scotophilus (yellow bats), family Vespertilionidae, diffuse in Bangladesh, India, Indonesia, Malaysia, Pakistan, Philippines, Sri Lanka, and Taiwan, reported as Nipah virus carrier. Furthermore, in China, the prevalence of anti- NiV or closely related virus antibodies was especially prominent among Daubenton’s bat (Mytotis daubentoni) and Rickett’s big-footed bat (Mytotis ricketti), two species of insectivorous bats of the genus Myotis, family Vespertilionidae. Daubenton’s bat (Mytotis daubentoni) is widely distributed throughout Britain, Europe, and as far as Japan and Korea. The presence of the Rickett’s big-footed bat (Mytotis ricketti) is limited to in China and Laos. A relatively high prevalence of antiHenipavirus antibody was also found in China among Leschenault’s Rousette fruit bat (Rousettus leschenaultia) of genus Rousettus, and in Ghana in the straw-coloured fruit bat (Eidolon helvum) of genus Eidolon, both of the family Pteropodidae.
_
A study was performed on free-ranging European insectivorous bats to assess the presence of paramyxovirus infection in these animals. The study involved 120 deceased bats of 15 different European species. Bayesian reconstruction of phylogenetic trees was performed in concordance with the current proposals of Paramyxoviridae taxonomy. Interestingly, the phylogenetic analysis confirmed the presence of the first three paramyxoviruses in European insectivorous bats. The genetic distance between these three novel paramyxoviruses and the closest related member resulted higher than that observed in other members within the paramyxovirus genera. This data suggested that all three viruses might be considered as new paramyxoviruses. Since, infected bats were found in close proximity to heavily populated human areas, a potential risk for a zoonotic paramyxovirus infection in Europe cannot be excluded. Recently, the occurrence of Henipaviruses in fruit bat populations in the north of Australia was explored. In particular, these authors evaluated the possibility that NiV were restricted to the west of Wallace’s Line. This line represents the biogeographic barrier existing between the Australo-Papuan and Wallacean region on the one hand, and Southeast Asia on the other, with different distribution of vertebrates and invertebrates. Data from this study demonstrated the presence of Nipah virus in both P. vampyrus and Rousettus amplexicaudatus the fruit bat populations localized on the eastern side of Wallace’s Line.
_
Amplifying host:
As previously stated, zoonotic diseases are spread from animals to humans via a reservoir host. However there can also be an intermediate host, in which the virus amplifies itself. However this intermediate host is not immune to the disease. Pigs have been identified as an intermediate and an amplifying host. An amplifying host is defined as a host in which the pathogen can become so prevalent to make it infectious. Pigs transmit NiV differently than bats in that they shed the virus primarily by coughing and the expulsion of respiratory secretions and saliva. It has been proved that the human outbreaks in Malaysia and Singapore with encephalitis and ~40 % case fatality occurred after contact with infected swine. The outbreak in Singapore ended when the importation of pigs from Malaysia was prohibited, and the outbreak in Malaysia ceased when over 1 million pigs (45 % of all pigs in Malaysia) were culled from the outbreak area and immediately surrounding areas. Serological studies indicated that NiV occurred in dogs, cats, horses and goats in the outbreak areas of Malaysia but all were effectively dead-end hosts. Apart from pigs, experimental infection has been produced in cats, golden hamsters and squirrel monkeys (Saimiri sciureus). Experimental studies have shown that infected cats shed NiV through the nasopharynx and in urine during viremic phase and of 2 cats infected, one recovered from experimental NiV infection with a high neutralizing antibody titer within 21 days. In a pregnant cat experimentally infected with NiV, the placenta and uterine fluid contained high levels of recoverable virus. The high levels of viral shedding combined with viral replication in fetal tissues suggested that both vertical and horizontal transmission of NiV could play a role in spillover events, an essential element in the epidemiology of Henipavirus infection.
_
In the course of investigating the origins of Nipah virus, ubiquitous peridomestic species were also extensively surveyed. The uniformly negative serology results from surveyed peridomestic rodents, insectivores, and birds in Malaysia (Asiah et al) indicate that these animals did not play a role as secondary reservoirs for Nipah virus. While evidence suggests that dogs readily acquired infection following close association with infected pigs, targeted surveillance indicated that Nipah virus did not spread horizontally within dog populations. Dogs in the outbreak area have also been shown to suffer from NiV infection. NiV infection was confirmed by immunohistochemical examination of one dead and one dying dog from the epidemic area in Malaysia. Both showed histologic evidence of severe disease. Many dogs sampled from the Bukit Pelanduk and Sepang epidemic area around the time of the pig culling had antibodies to NiV. However, Mills et al. analysed the serologic status in the dogs carefully and concluded that in the absence of infected pigs, dogs were not a secondary reservoir for NiV.
______
______
Nipah virus infections in pigs:
The incubation period in pigs is estimated to be 7 to 14 days, but it may be as short as four days. Experimentally infected cats developed clinical signs after 6-8 days and experimentally infected ferrets after 6-10 days.
Pathogenesis:
NiV targets the vascular, nervous, and lymphoreticular systems. Once NiV enters the oronasal cavity, the virions infect epithelial cells, immune cells, and peripheral nerve endings of cranial nerves. NiV can then directly follow nerves into the brain in some pigs and cause neurological symptoms. NiV also targets endothelial cells of small vessels as well as immune cells, leading to widespread viral dissemination. Viremia results in increased viral load in tissue parenchyma, including the central nervous system and the lower respiratory tract. NiV infects monocytes and T lymphocytes, and NiV antigen has been found in macrophages and dendritic cells. Immune cell depletion, including T lymphocytes, may play a role in increased risk of secondary infections.
Clinical Signs:
Young piglets are most likely to die from NiV. Sucking piglets may exhibit leg weakness, muscle tremors, and neurological twitches. In growing pigs, NiV infection may be asymptomatic or cause acute, febrile disease with respiratory and CNS signs. A characteristic respiratory sign in affected pigs is a “barking” cough, although there may only be labored to open-mouth breathing. CNS signs associated with disease include muscle fasciculation, leg weakness, ataxia, and spastic paresis. Dead pigs may present with a bloody nasal discharge.
Mortality is not common in sows or boars affected by NiV, although adults can die suddenly with no symptoms or within 24 hours after the onset of symptoms. Neurological signs seen in adults include tongue drooping, frothy salivation, head pressing, agitation, tetanic spasms and seizures, and pharyngeal muscle paralysis. Bloody nasal discharge can occur post mortem, and abortions have also been observed.
Post mortem Lesions:
Gross lesions include pulmonary consolidation and distended interlobular septa on cut surfaces. Frothy exudate, sometimes laced with blood, can fill the bronchi and trachea of affected pigs. Enlarged bronchial, submandibular, and mesenteric lymph nodes were commonly found. Lesions associated with neurological symptoms include meningeal congestion and edema. Meningitis or meningoencephalitis was more frequently seen in neurological cases than was encephalitis. Microscopic lesions seen in affected pigs include giant-cell pneumonia with multinucleated syncytia in the respiratory epithelium. Syncytia can also be found in endothelial cells of small blood and lymph vessels. Some affected pigs presented with meningeal inflammatory infiltrate. Lymphangitis may also be seen coupled with lymphocyte necrosis and depletion.
Nipah virus was widespread in pigs during the 1998-1999 outbreak in Malaysia. Before this virus was eradicated from domesticated swine, seropositive animals were found on approximately 5.6% of all pig farms. On one farm, more than 95% of all sows and 90% of the piglets had antibodies to this virus. The morbidity rate is estimated to approach 70-100%, but the mortality rate is low (e.g., 1-5% in 1-6 month old pigs) except in piglets. Mortality in the latter age group was approximately 40% in Malaysia, although neglect of the piglets by sick sows may have also played a role.
Nipah virus infection in other species:
Horses thought to be infected with Nipah virus in the Philippines either developed acute, fatal neurological signs or died suddenly with no apparent preceding illness. Although significant numbers of dogs and cats may have been infected on farms in Malaysia, clinical cases have been published for only two dogs. One of these animals had died of the illness, and the clinical signs were not described. In the other dog, the disease resembled canine distemper; the clinical signs included fever, respiratory distress, conjunctivitis, and mucopurulent nasal and conjunctival discharges. Experimental inoculation of cats with Nipah virus resulted in severe respiratory signs with fever, depression, an increased respiratory rate and dyspnea. Three cats thought to have been infected during an outbreak in the Philippines were found dead, while a fourth was moribund with terminal bleeding from the nose and mouth. Experimentally infected ferrets developed severe depression, serous nasal discharge, coughing, dyspnea, tremors and hindlimb paresis. Neurological and/or respiratory signs, which can be severe, have also been reported in some experimentally infected hamsters. An unproductive cough, poor growth, severe respiratory signs and deaths were documented in naturally infected goats in Malaysia. Two goats associated with a Nipah case in Bangladesh had a febrile neurological syndrome, but whether their illness was caused by Nipah virus or another disease is unknown. Infections in fruit bats appear to be asymptomatic.
_____
_____
Transmission and spread of nipah virus disease:
NiV infection has been found in 5 of 14 fruit bat species, with the highest being Pteropus hypomelanus with a 31% infection rate. Despite having a relatively high infection rate, the species of Pteropus fruit bats are not susceptible to the virus. In Pteropus bats, Nipah virus has been found repeatedly in urine, and viral RNA has been detected rarely in oropharyngeal swabs and rectal swabs from naturally or experimentally infected bats. It has also been found in fruit that had been partially eaten by bats. Despite high seroprevalence rates, only a few bats in a colony may shed the virus at any given time, and excretion from the colony may be sporadic. How bats transmit this virus to domesticated animals is uncertain, but ingestion of contaminated fruit, water, or aborted bat fetuses or birth products (e.g., by pigs) is suspected. Nipah virus is highly contagious in swine, which can act as amplifying hosts and shed this virus in respiratory secretions and saliva. Experimental infection in pigs show that NiV is detectable in oropharyngeal and nasal swabs as early as two days post-infection and experimentally infected animals continued to shed virus until three weeks post-infection.
_
Sources of virus:
- Nipah virus has been found in urine and uterine fluids of wild pteropid bats; experimentally isolated from urine, kidney and uterus of infected bats
- Virus may be found in fruit contaminated with bat saliva or urine
- Other logical sources for infection are contaminated drinking water and access to aborted bat foetuses or other fluids/tissues of parturition
- Once infected, pigs shed Nipah virus in respiratory secretions and saliva; urine of infected swine should also be considered a risk
- Role of other animals as a source of virus in outbreaks is less clear though virus has been isolated from feline respiratory secretions, urine, the placenta and embryonic fluids
- Sources of infective material for humans include swine respiratory secretions or via fruit or juice contaminated by bat secretions (e.g. unpasteurised date palms or juice)
_
Infected bats shed virus in their excretion and secretion such as saliva, urine, semen and excreta but they are symptomless carriers. NiV is highly contagious among swine populations and spreads via direct contact, exposure to infected secretions by coughing, and possibly aerosolization. Experimentally infected pigs have shown ability to excrete NiV as early as 4 days post-infection from the oropharynx, and NiV can also be shed in nasal secretions. Direct contact with infected pigs was identified as the predominant mode of transmission in humans when it was first recognized in a large outbreak in Malaysia in 1999. Ninety percent of the infected people in the 1998-1999 outbreaks were pig farmers or had contact with pigs. Transmission of Nipah virus to humans may occur after direct contact with infected bats, infected pigs, or from other Nipah virus infected people. Two routes of transmission of Nipah virus have also been identified from its natural reservoir to human: drinking of raw date palm sap contaminated with NiV and close physical contact with Nipah infected patients. The person-to person transmission may occur from close physical contact, especially by contact with body fluids.
_
There is strong evidence that emergence of bat-related viral infection communicable to humans and animals have been attributed to the loss of natural habitats of bats. As the flying fox habitat is destroyed by human activity the bats get stressed and hungry, their immune system gets weaker, their virus load goes up and a lot of virus spills out in their urine and saliva. Similar fluctuation of virus shedding may be associated with the stressful physiological conditions or seasons. Evidence of seasonal preference of transmission in P. lylei was recently demonstrated in a study in Thailand. The period April-June was the time (highest in May) when viral RNA could be mainly detected in urine which was associated with a fluctuation of population numbers that was observed only in May and correlated with young bats leaving to fly. There were focal outbreaks of NiV in Bangladesh and India in 2001 during winter. Drinking of fresh date palm sap, possibly contaminated by fruit bats (P. giganteus) during the winter season, may have been responsible for indirect transmission of Nipah virus to humans. There is circumstantial evidence of human-to-human transmission in India in 2001. During the outbreak in Siliguri, 33 health workers and hospital visitors became ill after exposure to patients hospitalized with Nipah virus illness, suggesting nosocomial infection. During the Bangladesh outbreak the virus is suggested to have been transmitted either directly or indirectly from infected bats to humans. Strong evidence indicative of human- to-human transmission of NiV was found in Bangladesh in 2004.
_
The virus is excreted in bat urine. In the original Malaysian outbreak, pigs became infected, possibly through eating fruit which had been contaminated by bats or directly from bat urine. The virus was excreted in pig urine, saliva and respiratory secretions, which is how humans, principally pig farmers, their families and abattoir workers, become infected. Cats, dogs and other domestic animals can also be affected. In the Bangladesh outbreaks humans become ill with no associated disease in animals and so they are thought to have been infected directly from bats. Drinking date palm sap (juice) which had been contaminated by bat droppings or saliva has been implicated. In addition human-to-human transmission has been documented in Bangladesh. Nipah virus is an emerging zoonotic disease that has occasionally been transmitted from person to person. In Bangladesh family members provide most of the hands-on care to patients in public hospitals. They expose themselves to patient saliva (and risk of disease transmission) by sleeping in the same bed, sharing patient’s uneaten food, and receiving a cough or sneeze in the face.
_
Transmission of Nipah virus to humans may occur after direct contact with infected bats, infected pigs, or from other NiV infected people. In Malaysia and Singapore, humans were apparently infected with Nipah virus only through close contact with infected pigs. The NiV strain identified in this outbreak appeared to have been transmitted initially from bats to pigs, with subsequent spread within pig populations. Incidental human infections resulted after exposure to infected pigs. No occurrence of person-to-person transmission was reported in this outbreak. Conversely, person-to-person transmission of Nipah virus in Bangladesh and India is regularly reported. This is most commonly seen in the family and caregivers of Nipah virus-infected patients. Transmission also occurs from direct exposure to infected bats. A common example is consumption of raw date palm sap contaminated with infectious bat excretions.
_
![]()
_
During the initial outbreaks in Malaysia and Singapore, most human infections resulted from direct contact with sick pigs or their contaminated tissues. Transmission is thought to have occurred via respiratory droplets, contact with throat or nasal secretions from the pigs, or contact with the tissue of sick animals. In swine, vertical transmission across the placenta, by iatrogenic means and in semen has been suggested but not confirmed. While the outbreak in Malaysia had progressed from the natural host (fruit bats), to amplification host (livestock) and finally to humans, in Bangladesh no amplification host was needed. People were somehow being directly infected by fruit bats. In the Bangladesh and India outbreaks, consumption of fruits or fruit products (e.g. raw date palm sap) contaminated with urine or saliva from infected fruit bats was the most likely source of infection. Other people seem to have been infected while working in the trees. During the later outbreaks in Bangladesh and India, Nipah virus spread directly from human-to-human through close contact with people’s secretions and excretions. In Siliguri, India, transmission of the virus was also reported within a health-care setting, where 75% of cases occurred among hospital staff or visitors. From 2001 to date, around half of reported cases in Bangladesh were due to human-to human transmission by close contact. Most of these infections occurred due to a small number of human transmitters, including one linked to 22 other human cases. Such persons are reminiscent of “super spreaders” in other diseases, most recently SARS. Nipah cases tend to occur in a cluster or as an outbreak, although 18 % of cases in Bangladesh were isolated.
_
Studies from the known human outbreaks of henipavirus in Australia, Malaysia, Singapore, Bangladesh, India, and the Philippines, show that the virus can be transmitted to human by three different routes: 1) from bats to humans who come in contact with virus-contaminated material (e.g., date palm sap); 2) from intermediate hosts such as pigs and horses; and 3) from infected humans. There is also epidemiological evidence that companion animals (including dogs and cats) can be infected with these viruses and although they can in theory transmit viruses to humans, no evidence is yet found of their role in transmission.
_
Transmission of the Nipah virus occurs when:
- Humans consume fruits bitten by bats
- Humans get into direct contact with infected bats, infected pigs or infected people
- Humans come into contact with body fluids of animals or people infected with the virus
_
Risk factors and transmission of Hendra virus:
All reported human cases of Hendra virus infection have been associated with exposure to ill horses, most of which were also confirmed to have been infected with Hendra virus. The mode of transmission was attributed to exposure to horse respiratory droplets, but it appears that close and/or prolonged contact is necessary for transmission. After Hendra virus was first identified, subsequent surveillance among 296 other potential contacts failed to identify any other antibody positive individuals, suggesting that the threshold for infectivity is low (McCormack et al., 1999). There is no evidence for subclinical infection or human-to-human transmission of Hendra virus.
_
Risk factors and mechanisms of transmission of Nipah virus:
Given the epidemiologic differences between cases in Malaysia, Bangladesh, and India, it is apparent that various factors play a role in the transmission of Nipah virus, including close exposure to intermediate zoonotic hosts, indirect contact with infected pteropid bats or exposure to their body secretions, and person-to-person transmission. In Malaysia and Singapore, direct contact with pigs, especially activities involving close contact, was the primary source of human Nipah virus infection (Parashar et al., 2000). Zoonotic transmission from pigs to humans probably occurred through respiratory droplets, given that infected pigs demonstrate both upper and lower respiratory vasculitis and have been shown in experimental studies to infect one another through the oral and respiratory route (Hooper and Williamson, 2000; Mohd Nor et al., 2000). Zoonotic transmission also appears to occur via close handling of infected tissue, as seen in the cases of infected abattoir workers (Paton et al., 1999). Exposure to infected pigs accounted for most cases of Nipah virus in humans, but 8% of case patients stated had no direct contact with pigs (Parashar et al., 2000). Furthermore, no obvious zoonotic source of transmission has been found in any of the Bangladesh outbreaks. The observation that many infected individuals in Bangladesh were under 19 years of age and had no exposure to pigs or other animals, in contrast to the Malaysia outbreak has led to the hypothesis that infection might have occurred by indirect contact with fruit bats or their secretions. It was observed that in Goalanda, boys ate fruit collected from trees where fruit bats were presumably foraging (Anon., 2004).
Nipah virus has been isolated from urine and respiratory secretions of humans with Nipah virus infection during the Malaysia outbreaks, suggesting the possibility of person-to-person transmission. In Malaysia, no evidence of person-to-person transmission was found despite extensive searching; but several households in both the 2001 and 2004 outbreaks in Bangladesh exhibited family clustering of cases, suggesting that limited person-to-person transmission might have occurred (Hsu et al., 2004; Anon., 2004). Person-to-person transmission was strongly suspected during the recent outbreak in Faridpur District. Testing of high-risk health care workers with patient contact during the Malaysia outbreaks revealed no evidence of nosocomial transmission. During the Siliguri outbreak, encephalitis cases developed among some hospital staff several days after the admission of patients with Nipah virus encephalitis, suggesting possible nosocomial transmission, but specific exposures were not assessed in affected individuals. There is high use of standard and respiratory precautions in Malaysia, but only about 40% of health care workers in Bangladesh during the outbreaks used any type of barrier precaution. These findings taken together suggest that person-to-person transmission of Nipah virus can occur, but that the transmission is rather inefficient, and probably requires prolonged close contact.
______
NiV Transmission from Bats to People:
The primary pathways of transmission from bats to people in Bangladesh are through contamination of raw date palm sap by bats with subsequent consumption by humans and through infection of domestic animals (cattle, pigs, and goats), presumably from consumption of food contaminated with bat saliva or urine with subsequent transmission to people. Approximately one-half of recognized Nipah case patients in Bangladesh developed their disease following person-to-person transmission of the virus. Efforts to prevent transmission should focus on decreasing bat access to date palm sap and reducing family members’ and friends’ exposure to infected patients’ saliva.
_
The exact mechanism of henipavirus transmission from bats to humans is poorly understood. Experimental studies in this area are severely hampered by 2 practical difficulties. First, because bats are wild animals, it is not easy to obtain relevant species of bats that have a known history of being free from prior infection with henipavirus or closely related viruses. Second, both HeV and NiV are biosafety level (BSL) 4 agents, and any infection studies have to be conducted in BSL4 facilities by skilled, highly trained operators. Nevertheless, on the basis of existing field and laboratory data, 3 hypotheses have been postulated. One is that masticated pellets of virus-contaminated residual fruit pulp spat out by flying foxes are consumed by susceptible animals, such as horses or pigs. The second is that urine from infected animals contaminates pastures or pig sties. The third is that infected fetal tissues or fluids contaminate pastures or sties and are ingested.
_
Epidemiological investigations in Bangladesh have identified three pathways of transmission of NiV from bats to people.
- The most frequently implicated route is ingestion of fresh date palm sap. Date palm sap is harvested from December through March, particularly in west central Bangladesh. A tap is cut into the tree trunk and sap flows slowly overnight into an open clay pot. Infrared camera studies confirm that P. giganteus bats frequently visit date palm sap trees and lick the sap during collection (Khan et al., 2008). NiV can survive for days on sugar-rich solutions such as fruit pulp (Fogarty et al., 2008). Most date palm sap is processed at high temperature to make molasses, but some is enjoyed as a fresh juice, drunk raw within a few hours of collection. In the 2005 Nipah outbreak in Tangail District, Bangladesh, the only exposure significantly associated with illness was drinking raw date palm sap (64% of case patients vs. 18% of control patients; odds ratio [OR], 7.9; 95% confidence interval [CI], 1.6–38; P = .01) (Luby et al., 2006). Twenty-one of the 23 index NiV case patients recognized in Bangladesh developed their initial symptoms during the December through March date palm sap collection season (Luby et al., 2009).
_
Fruit bats (P. giganteus) are a nuisance to date palm sap collectors because the bats drink the sap at night from the clay pots used to collect the sap. The investigations by Luby et al. suggested that NiV was transmitted from P. giganteus to persons through drinking fresh date palm sap. Date palm sap is a national delicacy that is enjoyed by millions of Bangladeshis each winter. However, the sap is occasionally contaminated with NiV-infected bat urine or saliva that contains a sufficient dose of NiV to be fatal to humans. In India, in a bat sample survey, NiV RNA was detected in a liver homogenate of P. giganteus captured in Myanaguri, West Bengal.
_
Collection of date palm sap:

The date palm trees are scratched to collect sap through a bamboo channel. An earthen pot is tied for collection of the sap. In Bangladesh, date palm sap has been identified as the most relevant risk factor related with the epidemiology of Nipah virus. In this country, it is very popular, used to make products like molasses, used as a sweetener in traditional cakes and desserts, and often consumed raw. Date palm sap is collected during the coolest months of the year, typically mid-December through Mid-February when humidity and temperatures permit efficient sap collection. Harvesters, known as gachis in Bangladesh, collect sap by cutting a v-shaped gouge into a date palm tree and hanging a container overnight.
_
In January 2005 in Bangladesh, an outbreak of Nipah virus infection in 12 people, 11 of whom died, was traced to drinking raw date-palm juice. Fruit bats, a nuisance to date-palm juice collectors, drink date-palm juice directly from the cut in the tree or the clay pot used to collect the sweet sap overnight. The juice is gathered in the morning and sold fresh as it ferments quickly and loses its sweet taste. Because palm juice is consumed within a few hours of harvest, Nipah virus, introduced into the juice by the fruit bats, might be able to survive in sufficient numbers for transmission.
_
- A second route of transmission for NiV from bats to people in Bangladesh is via domestic animals. Fruit bats commonly drop partially eaten saliva-laden fruit. Domestic animals in Bangladesh forage for such food. Date palm sap that is contaminated with bat feces and so is unfit for human consumption is also occasionally fed to domestic animals. The domestic animals may become infected with NiV, and shed the virus to other animals, including humans. Contact with a sick cow in Meherpur, Bangladesh in 2001 was strongly associated with Nipah infection (OR, 7.9; 95% CI, 2.2–27.7; P = .001) (Hsu et al., 2004). A pig herd visited the community two weeks before the 2003 Nipah outbreak in Naogaon and contact with the pigs was associated with illness (OR, 6.1; 95% CI, 1.3–27.8; P = .007) (Khan et al., 2008). In 2004, one family explained that they owned 2 goats that their son frequently played with. The goats became ill with fever, difficulty walking, walking in circles, and frothing at the mouth. The parents believe their son had contact with goat saliva while the goats were ill. Both goats died. Within 2 weeks of the goats’ death, the child developed encephalitis that was confirmed to be Nipah by antibody testing. Several outbreaks of NiV have been reported in human beings in Bangladesh from 2001 to 2013 and three outbreaks in India, but none had showed any involvement of pigs.
_
- Third, some people may come into direct contact with NiV-infected bat secretions. In the Goalando outbreak in 2004, persons who climbed trees were more likely to develop NiV infection than were control patients (OR, 8.2; 95% CI, 1.3–∞) (Montgomery et al., 2008).
_
Convergence of Humans, Bats, Trees, and Culture in Nipah Virus Transmission, Bangladesh, a 2017 study:
Preventing emergence of new zoonotic viruses depends on understanding determinants for human risk. Nipah virus (NiV) is a lethal zoonotic pathogen that has spilled over from bats into human populations, with limited person-to-person transmission. Authors examined ecologic and human behavioral drivers of geographic variation for risk of NiV infection in Bangladesh. They visited 60 villages during 2011–2013 where cases of infection with NiV were identified and 147 control villages. They compared case villages with control villages for most likely drivers for risk of infection, including number of bats, persons, and date palm sap trees, and human date palm sap consumption behavior. Case villages were similar to control villages in many ways, including number of bats, persons, and date palm sap trees, but had a higher proportion of households in which someone drank sap. Reducing human consumption of sap could reduce virus transmission and risk for emergence of a more highly transmissible NiV strain.
_
![]()
Figure above shows zoonotic transmission cycles of Nipah virus in Malaysia and Bangladesh. Pteropus species fruit bats are the natural reservoir of Nipah virus. In Malaysia (left), Nipah virus was transmitted from bats roosting in fruit trees on pig farms to pigs; subsequently, pigs transmitted Nipah virus to people in close contact with the pigs. In Bangladesh (right), Nipah virus is thought to be transmitted via the consumption of raw date palm sap. While date palm sap is collected, bats drink from the sap stream or collection pots and contaminate the sap with Nipah virus through their saliva or urine. People become infected with Nipah virus after drinking this contaminated date palm sap. Subsequently, these infected people can transmit Nipah virus to others via close contact.
_______
Hamster study:
Because of the heavy prevalence of virus transmission via respiratory secretions it is assumed the main site of replication occurs in the tonsils of an infected host. This is supported by research done in the transmission of NiV in hamsters. de Wit et al (2011) observed viral shedding of hamsters inoculated with three different doses of varied strength of NiV as seen in the figure below. The researchers then examined varied forms of shedding of the virus nasally, oropharyngeally, urogenitally, and rectally. The results show, regardless of the dosing, all three dosages resulted in positive viral loads in the oropharyngeal region as soon as two days post inoculation.
![]()
Figure above shows Data collected in observing the shedding of Nipah virus RNA in inoculated hamsters. Black bars indicate smallest dosage, grey bars was the middle range dosage, white bars was the highest dosage. All differences from days post inoculation were significant. Swabs were taken oropharyngenally.
_____
_____
Person to person transmission:
Several Bangladesh Nipah outbreaks resulted from person-to-person transmission. The clearest illustration of person-to-person NiV transmission occurred during the Faridpur outbreak in 2004 (Gurley et al., 2007). Four persons who cared for the index patient—his mother, his son, his aunt, and a neighbor—became ill 15–27 days after the index patient first developed illness. During her hospitalization, the index patient’s aunt was cared for by a popular religious leader who lived in a nearby village and who became ill 13 days later. When the religious leader became seriously ill, many of his relatives and members of his religious community visited at his home. Twenty-two persons developed Nipah infection after contact with the religious leader. One of these followers moved to his family’s house in an adjacent village to receive care after becoming ill where he was cared for by a friend and 2 family members. These 3 caregivers and a rickshaw driver, who helped carry him to the hospital as his condition deteriorated, became ill. Ultimately, the chain of transmission involved 5 generations and affected 34 people (Gurley et al., 2007). Physical contact with an NiV-infected patient who later died (OR, 13.4; 95% CI, 2.0–89) was the strongest risk factor for developing NiV infection in the outbreak.
_
The transmission pattern in Faridpur is not unique. For example, in 2007 in Thakurgaon, 6 family members and friends who cared for an NiV-infected patient developed Nipah infection. Case patients were more likely than control patients to have been in the same room when the index case was coughing (ICDDRB, 2007). In a review of the 122 Nipah case patients identified in Bangladesh from 2001 through 2007, 62 (51%) developed illness after close contact with another Nipah patient (Luby et al., 2009). A small minority of patients infected with NiV (i.e., 9 [7%] of 122 recognized cases) transmitted NiV to 62 other persons.
_
Respiratory secretions appear to be particularly important for person-to- person transmission of NiV. NiV RNA is readily identified in the saliva of infected patients (Chua et al., 2001; Harcourt et al., 2005). Anthropological investigations during the Faridpur outbreak highlighted multiple opportunities for the transfer of NiV contaminated saliva from a sick patient to care providers (Blum et al., 2009). Social norms in Bangladesh require family members to maintain close physical contact during illness. The more severe the illness, the more hands-on care is expected. Family members and friends without formal health care or infection control training provided nearly all the hands on care to Nipah patients both at home and in the hospital (Hadley et al., 2007). Care providers during the Faridpur outbreak continued to share eating utensils and drinking glasses with sick patients. Leftovers of food offered to ill Nipah patients were commonly distributed to other family members. Family members maintained their regular sleeping arrangements, which often involved sleeping in the same bed with a sick, coughing Nipah patient. There was a particularly strong desire to have close physical contact near the time of death, demonstrated by such behaviors as cradling the patients head on the family member’s lap, attempting to give liquids to the patient with a spoon or glass between bouts of coughing, or hugging and kissing the sick patient (Blum et al., 2009). In both the Faridpur outbreak in 2004 and the Thakurgaon outbreak in 2007, persons who were in a room when a Nipah patient was coughing or sneezing were at increased risk of Nipah virus infection (Gurley et al., 2007a; ICDDRB, 2007). Across all recognized outbreaks in Bangladesh from 2001 through 2007, Nipah patients with respiratory symptoms were more likely to transmit Nipah (Luby et al., 2009).
_
The capacity for NiV to spread in hospital settings to both staff and patients was clearly illustrated in a large outbreak affecting 66 people in Siliguri, India in 2001. The outbreak apparently originated from an unidentified patient admitted to Siliguri District Hospital who transmitted infection to 11 additional patients, all of whom were transferred to other facilities. In 2 of the facilities, subsequent transmission infected 25 staff and 8 visitors (Chadha et al., 2006). However, transmission to health care workers is rarely recognized. Among a cohort of 338 health care workers who cared for Nipah patients at 3 Malaysian hospitals and reported a combined 89 episodes of Nipah patient blood or body fluid directly contacting their bare skin, 39 splash exposures of blood or body fluid into their eyes, nose or mouth, and 12 needle stick injuries, none developed clinical illness associated with Nipah infection (Mounts et al., 2001). Health care workers in Bangladesh have much less direct physical contact with patients than in western hospitals (Hadley et al., 2007). Hands-on care is generally provided by family members and friends. No health care workers in Bangladesh who cared for identified Nipah patients have been identified with illness, although confirmed cases include 1 physician whose source of infection is unknown. A serosurvey among 105 health care workers who cared for at least 1 of 7 patients admitted with Nipah infection at one hospital in Bangladesh identified 2 health care workers with serological evidence of NiV infection; however, their antibody responses (IgG only, no IgM) and lack of symptoms suggest a previous infection, not recent nosocomial transmission (Gurley et al., 2007).
_
Might person-to-person transmission be associated with particular strains of NiV that have genetic characteristics that lead to person-to-person transmission? The closely related strains in Malaysia resulted in less frequent and less severe respiratory disease than observed in Bangladesh and were not associated with frequent person-to-person transmission. However, the pattern of the outbreaks in Bangladesh and India suggests that person-to-person transmission is more dependent on the characteristics of the occasional Nipah transmitter than a specific strain. If the NiV strain was central to person-to-person transmission, then secondary cases of NiV would be more likely to become NiV transmitters, than primary cases (because secondary cases would already have selected for strains predisposed to person-to-person transmission). However, in the review of 7 years of human Nipah infection in Bangladesh, secondary cases were no more or less likely to become Nipah transmitters than were primary cases (Luby et al., 2009). All persons who transmitted Nipah died, suggesting that late stages of infection, presumably with high virus titers, increases the risk of transmission. Even the pattern in Siliguri, the largest recognized Nipah outbreak from apparent person-to-person transmission, is consistent with the review of 7 years of human Nipah infection in Bangladesh. The unidentified index case in Siliguri District Hospital infected 11 patients, 2 of whom infected an additional 33 patients. The 13 day duration of the outbreak at Medinova Hospital suggests 2 generations of transmission likely occurred there. Taken together, this pattern suggests 4 NiV transmitters propagated human infection across 4 generations. There were 67 cases (66 recognized plus the unidentified index case), 4 (5.9%) of whom became Nipah transmitters, a proportion very close to the 7% recognized in Bangladesh. This suggests that the virus strain responsible for this largest recognized person-to-person outbreak was not exceptional. Its rate of secondary transmission was similar to other strains circulating in South Asia. Two outbreaks of encephalitis, both caused by Nipah viruses, occurred in separate areas of western Bangladesh in 2007. Each outbreak lasted <3 weeks and had high (43% and 63%) case-fatality ratios. In both the outbreaks cases were clustered in time and place with a secondary peak of cases following the initial cases suggesting person-to-person transmission. The capacity of the virus to spread from human to human increases the need for enhanced infection prevention measures in a densely populated country such as Bangladesh.
_
According to Gurley et al., 2007 there is significant evidence that person-to-person contact will cause infection. The person-to-person transmissibility factors include having (1) touched or received a cough or sneeze in the face, (2) any contact with someone who later died, was febrile, unconscious, or had respiratory difficulty, and (3) visited the home, and possibly, the village an infected person.
________
The Nature of Exposure drives Transmission of Nipah Viruses from Malaysia and Bangladesh, a 2016 study in Ferrets:
Nipah viruses cause outbreaks of severe human disease with high fatality rates. Different patterns of transmission have been associated with recurrent outbreaks caused by Nipah virus (NiV) in Bangladesh (BD), where person-to-person transmission is a major pathway for human infection, compared to an outbreak in Malaysia (MY), where pig-to-human transmission accounted for virtually all human infections. To date, there have been limited comparative studies that address the question of whether this difference may be attributed to differences between geographical isolates of NiV at the genome level, or to other factors in play during human outbreaks of disease. In a time course study, ferrets were oronasally exposed to equivalent doses of NiV-BD or NiV-MY. More rapid onset of productive infection and higher levels of virus replication in respiratory tract tissues were seen for NiV-BD compared to NiV-MY, corroborating author’s previous report of increased oral shedding of NiV-BD in ferrets and suggesting a contributory mechanism for increased NiV-BD transmission between people compared to NiV-MY. However, authors recognize that transmission occurs within a social and environmental framework that may have an important and differentiating role in NiV transmission rates. With this in mind, ferret-to-ferret transmission of NiV-BD and NiV-MY was assessed under differing viral exposure conditions. Transmission was not identified for either virus when naïve ferrets were cohoused with experimentally-infected animals. In contrast, all naïve ferrets developed acute infection following assisted and direct exposure to oronasal fluid from animals that were shedding either NiV-BD or NiV-MY. These findings for ferrets indicate that, although NiV-BD may be shed at higher levels than NiV-MY, transmission risk may be equivalently low under exposure conditions provided by cohabitation alone. In contrast, active transfer of infected bodily fluids consistently results in transmission, regardless of the virus strain. These observations suggest that the risk of NiV transmission is underpinned by social and environmental factors, and will have practical implications for managing transmission risk during outbreaks of human disease.
_
In earlier work conducted in the ferret, authors found significantly higher levels of viral RNA in oropharyngeal secretions of animals infected with NiV-BD by comparison with NiV-MY, suggesting that the two strains may differ in their replication efficiency at sites of potential relevance to transmission. Higher levels of oral shedding of the Bangladesh strain of Nipah virus might be a key factor in onward transmission in outbreaks among humans. The observations of a trend for earlier onset of respiratory tract shedding, a higher rate of respiratory tract shedding during clinical disease, and a greater magnitude of viral shedding from NiV-BD infected ferrets constitute a virus-dependent contribution to the basic reproduction number (R0) which might result in a difference in R0 between the two virus strains. However, viral factors are only one determinant of R0; social factors–for example, those which define the interactions between affected individuals and the other members of the population–also play an important role in the likelihood of transmission. Accordingly, authors adapted studies designed to assess the transmissibility of influenza virus strains in ferrets to the operational constraints of the BSL-4 environment and completed two small scale comparisons of NiV strain transmissibility, providing different contact exposure opportunities which were duplicated for each virus strain. Authors found in this study that although there may be higher levels of viral shedding with NiV-BD compared to NiV-MY, the attack rate for both viruses may be equivalently low under exposure conditions provided by cohabitation alone. In contrast, the attack rates for each virus are similarly high when there is active transfer of infected bodily fluids.
Findings from the studies presented herein are consistent with the view that the risk of human-to-human transmission of certain isolates of NiV is more plausibly underpinned by environmental factors in play with each outbreak event, than by inherent differences in tissue tropism or pathogenicity between NiV strains. In particular, authors have demonstrated that under experimental conditions that mimic high-risk human exposure events as reported in Bangladesh, NiV-BD and NiV-MY cause similar outcomes of infection in ferrets. Their observations suggest that a critical driver of onward NiV transmission is the nature of the interaction between subjects and that its impact may be irrespective of virus strain. In particular, the social and cultural contexts in which infection events occur, and how these influence the management of the typical infected individual, are the pivotal determinants of the likelihood of onward transmission.
______
______
Nipah spread by droplet contact transmission and not by airborne transmission:
A study showed that primary human basal respiratory epithelial cells are susceptible to both NiV-Malaysia (M) and -Bangladesh (B) strains causing robust pro-inflammatory responses, and that infection of small airways and not large airways results in disruption of the epithelium integrity and host responses recruiting human immune cells. While human-to-human transmission has been observed only in outbreaks with NiV-B, these data suggest that both NiV and HeV have the potential for human-to-human transmission through droplet contact transmission. Some diseases can be transferred by infected droplets contacting surfaces of the eye, nose, or mouth. This is referred to as droplet contact transmission. Droplets containing microorganisms can be generated when an infected person coughs, sneezes, or talks. Droplets are too large to be airborne for long periods of time, and quickly settle out of air. Droplet transmission can be reduced with the use of personal protective barriers, such as face masks and goggles. Frequent hand cleansing, especially with instant hand sanitizers, can help prevent droplet transmission. Measles and SARS are examples of diseases capable of droplet contact transmission. Maintain >1 full-stretched arm distance (1 metre or 3 feet) from patient to prevent droplet contact transmission.
The main difference between airborne and droplet contact transmission is in the size of infective particles. Particle less than 5 to 10 micron is airborne particles and larger particles for droplet contact transmission. Airborne transmission refers to situations where droplet nuclei (residue from evaporated droplets) or dust particles containing microorganisms can remain suspended in air for long periods of time. These organisms must be capable of surviving for long periods of time outside the body and must be resistant to drying. Droplet nuclei can travel on air currents over considerable distances. Even after infected person has left room, room air still contains infectious droplet nuclei capable of transmission. Many common infections can spread by airborne transmission at least in some cases, including: Anthrax (inhalational), Chickenpox, Influenza, Measles, Smallpox, Cryptococcosis, and Tuberculosis.
______
Surface-to-person transmission:
Humans have been found to be infected by three different pathways, Pteropus fruit bat to human, pig to human, and human to human. Most recent transmission of NiV has been a result of human to human transmission through close contact through respiratory secretions or urine. Although not completely understood, the molecular mechanism of Nipah virus transmission is hypothesized to be by fomites. Fomites are non-living objects which are able to carry infectious material like vectors. Currently this is the main hypothesis because of the increased number of NiV infections being picked up in hospitals. It is unknown how long the virus can survive outside a host. Data collected from various hospitals in Bangladesh where patients infected with NiV were treated showed a high amount of NiV RNA on hospital surfaces. Transmission of NiV through fomites is plausible. Many paramyxoviruses, including respiratory syncytial virus, parainfluenza viruses 1–4, and human metapneumoviruses, have been identified on hospital surfaces, and fomiteborne transmission of these pathogens has been reported. Past studies have indicated that other paramyxoviruses can survive on surfaces for up to 10 hours and be a source of infection for patients, healthcare workers, and hospital visitors. Although it is not known how long NiV remains infectious in the environment, researchers hypothesize that surfaces might play an important role in NiV transmission for several reasons: hospital surfaces in Bangladesh are not routinely cleaned; new patients frequently use the same bed sheets used by previous occupants; caregivers and healthcare workers frequently come into contact with contaminated surfaces; and hand washing by caregivers and healthcare workers occurs infrequently because of several barriers, including a lack of running water in hospitals.
_
Nipah virus can’t survive in fruits: say scientists from National Institute of Virology Pune India:
“Viruses are obligatory parasites. They cannot multiply in fruits. Like any other virus, Nipah virus needs animal or human cells to survive and multiply,” National Institute of Virology (NIV) director and senior scientist Devendra Mourya said. India’s fruit exports dipped after the outbreak of Nipah in Kerala. The deadly virus claimed 17 lives in the southern state, following which India’s fruit exports and consumption within the country went down significantly. The scientists urged citizens not to shy away from eating fruits because they did not provide the virus with the mechanism to survive and spread. Nipah virus is spread by fruit bats (Pteropus giganteus). A very small proportion of these bats secrete the virus. Human beings rarely come into contact of the infected secretion and develop the disease. The virus may infect a human through fruits only when an individual consumes a fruit nibbled by an infected bat immediately after the fruit came in contact with its infected saliva. In this case, the virus which has got on to the fruit surface through its saliva gets human cells to survive. Any time gap in between kills the virus then and there. The virus cannot survive in fruits.
______
My view:
Virus is a link between living and non-living matter. Although virus is an obligate intra-cellular parasite, virus outside living cells exist as particles which can be viable or non-viable. Non-viable virus particle in the environment is dead, cannot enter in host cell (e.g. empty capsids lacking any genetic material). Viable virus particle in environment is a state between living and non-living matter. Such particles can survive for hours to days depending on environmental conditions such as temperature, pH and chemical milieu. They can enter host cell and they are infectious or non-infectious. Infectious means completion of the replication cycle, from entry to synthesis of new particles in the host cell independently. In example below, it specifically means PFU, but in fact many viruses cannot form plaques on any cells. Non-infectious viable virus particle means it is unable to replicate in host cell on its own due to variety of reasons although it can enter host cell. Non-infectious virus particle can replicate with assistance. Smallpox virus can remain infectious for long periods even when outside of a host while Ebola virus left alone on a surface will begin to disintegrate rather quickly. Airborne transmission of Chickenpox, Influenza, Measles and Smallpox proves that these viruses can survive in air for long period of time outside body. How long Nipah virus can remain viable in the general environment is uncertain; however, it can survive for up to 3 days in some fruit juices or mango fruit, and for at least 7 days in artificial date palm sap (13% sucrose and 0.21% bovine serum albumin in water, pH 7.0) held at 22°C. NiV can survive for days on sugar-rich solutions such as fruit pulp (Fogarty et al., 2008). This virus is reported to have a half-life of 18 hours in the urine of fruit bats.
_
In virology, a plaque-forming unit (PFU) is a measure of the number of particles capable of forming plaques per unit volume, such as virus particles. It is a functional measurement rather than a measurement of the absolute quantity of particles: viral particles that are defective or which fail to infect their target cell will not produce a plaque and thus will not be counted. A plaque is interpreted as the result of a single independently infectious particle. The term particle-to-PFU ratio refers to the number of viral particles required to form one plaque in a plaque assay. It is a measure of the efficiency by which a virus infects cultured cells. It is a measure of the fraction of virus particles in a given sample that can complete an infectious cycle. For many bacteriophages, the particle-to-PFU ratio approaches 1, which is the lowest value that can be obtained. A value of 1 means that every virus particle in the sample is able to form a plaque.
For animal viruses, the particle-to-PFU ratio is often much higher, from 1 to 1000. These high values complicate the study of animal viruses. When the particle-to-PFU ratio is high, one can never be certain that properties measured in infected cells are those of the infectious or the non-infectious viral particles. The linear nature of the dose-response curve indicates that a single virion is capable of initiating an infection. However, the high particle-to-PFU ratio of many viruses shows that not all virions are successful. A high particle-to-PFU ratio is sometimes caused by the presence of noninfectious particles with genomes that harbor lethal mutations or that have been damaged during growth or purification. Another explanation is that although all viruses in a preparation are in fact capable of initiating infection, not all of them succeed because of the complexity of the infectious cycle. Failure at any one step in the cycle prevents completion. A high particle-to-PFU ratio does not indicate that most particles are defective, but that they failed to complete the infection. RNA viruses may have high particle-to-PFU ratios since these viruses exist as quasi species due to RNA-dependent RNA polymerase associated mistakes (as many as 10−3 – 10−5 mutations per nucleotide per replication event).
_
The synopsis of virus viability in environment is depicted below:
_____
Henipavirus susceptibility to environmental variables, a 2007 study:
The purpose of this study was to measure the persistence of henipaviruses under various environmental conditions and thereby gain an insight into likely mechanisms of transmission. Henipaviruses survived for more than four days at 22°C in pH-neutral fruit bat urine but were sensitive to higher temperatures and pH changes. On mango flesh, survival time varied depending on temperature and fruit pH, ranging from two hours to more than two days. Desiccation of viruses substantially reduced survival time to less than two hours. The conditions under which date palm sap is collected and consumed provide an opportunity for NiV transmission. The sap is collected at night during the coldest months of the year when the average minimum temperature is 13°C −15°C. The pH of palm sap is near neutral and it is consumed within a few hours after sunrise before it ferments. While date palm sap was not available for testing in this study, under these conditions, the current study indicates that there is likely to be only minimal loss of viral titre before consumption.
______
Transmission of NiV in hospital settings:
Transmission of NiV in hospital settings was first identified in 2001 during an outbreak in Siliguri, India, and in several outbreaks in Bangladesh since 2004. In the outbreak in Siliguri, 66 persons were infected, and of the 60 for whom exposure was known, 45 (75%) acquired infection during their hospital stay (11 patients admitted for other illness, 25 hospital staff, and 8 persons who visited an infected patient). In Bangladesh, during the 2010–2011 Nipah outbreak, 2 hospital staff (1 physician, 1 hospital cleaner) were infected. NiV RNA has repeatedly been identified in infected patients’ oral secretions, and epidemiologic evidence suggests that exposure to respiratory secretions is a likely route of NiV transmission from patient to caregiver. In 2004, during an NiV outbreak with person-to-person transmission in Bangladesh, NiV RNA was found on a hospital wall near where an NiV patient received care.
_
Hospital staff and family caregivers can acquire infections through direct patient contact or contaminated fomites. Healthcare workers (i.e., doctors and nurses) have direct contact with patients; other staff, such as hospital cleaners, and visitors, who are not involved in patient care, might have contact only with hospital surfaces. Possible contamination of nearby hospital surfaces by Nipah patients with infectious bodily secretions, coupled with a lack of infection control measures in low-income hospitals, puts healthcare workers, caregivers, visitors, and other patients in the ward at risk for NiV infection by contaminated hospital surfaces. Propagation of a highly fatal pathogen with the capacity for person-to-person transmission within resource-constrained healthcare settings increases the risk for broader outbreaks.
_
Nipah Virus Contamination of Hospital Surfaces during Outbreaks in Bangladesh between 2013–2014, a 2018 study:
Nipah virus (NiV) has been transmitted from patient to caregivers in Bangladesh presumably through oral secretions. Authors aimed to detect whether NiV-infected patients contaminate hospital surfaces with the virus. During December 2013–April 2014, they collected 1 swab sample from 5 surfaces near NiV-infected patients and tested surface and oral swab samples by real-time reverse transcription PCR for NiV RNA. They identified 16 Nipah patients; 12 cases were laboratory-confirmed and 4 probable. Of the 12 laboratory-confirmed cases, 10 showed NiV RNA in oral swab specimens. They obtained surface swab samples for 6 Nipah patients; 5 had evidence of NiV RNA on >1 surface: 4 patients contaminated towels, 3 bed sheets, and 1 the bed rail. Patients with NiV RNA in oral swab samples were significantly more likely than other Nipah patients to die. To reduce the risk for fomite transmission of NiV, infection control should target hospital surfaces.
______
______
Nipah virus transmission in animal models:
Since data from human cases are sparse, this knowledge is largely based on the results of studies performed in animal models that recapitulate Nipah virus disease in humans.
Nipah virus transmission in its natural and intermediate hosts:
How Nipah virus is transmitted between bats is unknown, and this question has thus far not been addressed experimentally. In fact, data on naturally as well as experimentally infected bats are sparse. Nipah virus is shed primarily via urine and also via saliva. This detection of Nipah virus in the urine and saliva of fruit bats and from partially eaten fruits, together with large colony sizes and abundant social interactions, suggest that direct contact transmission as well as fomite transmission are important routes of transmission within the natural host species.
It is obvious from the epidemiology of the Nipah virus outbreak in Malaysia and Singapore, with the virus spreading quickly across farms and states with the movement of infected pigs, that pigs are very susceptible to, and efficient spreaders of Nipah virus. Transmission of Nipah virus in the intermediate host was not studied in detail experimentally; however, inoculated pigs transmitted Nipah virus to two naïve cage mates. One of the two contact pigs developed mild, transient signs of disease; both animals shed infectious virus and seroconverted.
_
Zoonotic transmission:
As discussed above, zoonotic transmission of Nipah virus occurred through an intermediate host in Malaysia and Singapore. Experimental inoculation of pigs resulted in virus shedding from the nose, throat and eyes; viral RNA as well as infectious virus could be detected. In contrast, detection of virus shedding was sporadic in experimentally inoculated flying foxes and isolation of virus from swabs or urine of naturally infected bats is rare. Thus, the role of pigs as an amplifying intermediate host, being in relatively close contact with humans during the outbreak in Malaysia and Singapore is clear.
There is epidemiological evidence that zoonotic transmission of Nipah virus during outbreaks in Bangladesh occurs mainly through the consumption of raw date palm sap. This date palm sap is harvested at night in Bangladesh during the winter months and bats have been observed drinking from the sap stream or the collection pot. It is thought that bats contaminate the date palm sap with their saliva, urine or faeces containing Nipah virus while drinking from it, and that people who subsequently drink the sap can acquire Nipah virus infection that way. This zoonotic transmission route was investigated experimentally in the Syrian hamster model. To experimentally investigate this epidemiological association between drinking of date palm sap and human cases of Nipah virus infection, researchers determined the viability of Nipah virus (strain Bangladesh/200401066) in artificial palm sap. At 22°C virus titers remained stable for at least 7 days, thus potentially allowing food-borne transmission. Next, researchers modeled food-borne Nipah virus infection by supplying Syrian hamsters with artificial palm sap containing Nipah virus. Drinking of 5×108 TCID50 of Nipah virus resulted in neurological disease in 5 out of 8 hamsters, indicating that food-borne transmission of Nipah virus can indeed occur. In comparison, intranasal (i.n.) inoculation with the same dose of Nipah virus resulted in lethal respiratory disease in all animals. In animals infected with Nipah virus via drinking, virus was detected in respiratory tissues rather than in the intestinal tract. Using fluorescently labelled Nipah virus particles, researchers showed that during drinking, a substantial amount of virus is deposited in the lungs, explaining the replication of Nipah virus in the respiratory tract of these hamsters. Virus replication was detected in the respiratory tract and in the CNS, but not in the intestinal tract, thus resembling the disease in humans in Bangladesh and providing experimental support for the food-borne zoonotic transmission of Nipah virus there. Besides the ability of Nipah virus to infect hamsters via the drinking route, Syrian hamsters infected via that route transmitted the virus through direct contact with naïve hamsters in 2 out of 24 transmission pairs.
Human-to-human transmission:
Nipah virus can be transmitted from infected patients to naïve individuals. During the outbreak in India in 2001, ∼75% of cases acquired the infection through nosocomial transmission. Human-to-human transmission seems to occur regularly during Nipah virus disease outbreaks in Bangladesh; for some of the outbreaks it was estimated that ∼50% of cases were the result of human-to-human transmission. Human-to-human transmission was modelled in the Syrian hamster to determine the route through which Nipah virus is transmitted. Upon intranasal inoculation of Syrian hamsters with Nipah virus, transmission to naïve hamsters was observed only in hamsters that were in direct contact, but not via fomites or aerosols. The animals that were infected through transmission did not develop signs of disease, but shedding of viral RNA was detected and animals developed a neutralizing antibody response against Nipah virus. Surprisingly, a Nipah virus strain isolated from a fatal case in Malaysia was transmitted as efficiently as a strain isolated from a fatal case in Bangladesh, indicating that the apparent absence of human-to-human transmission during the outbreak in Malaysia was most likely not due to a difference in the ability of the virus to transmit, but most likely due to different health-care practices and cultural differences, resulting in more contact between sick individuals and their family members.
_____
_____
There are many grey areas in nipah transmission and we need more research. Exposures not associated with NiV transmission is one of the grey areas:
Outbreak investigations have both identified important routes of transmission of human NiV infection, and identified exposures not associated with transmission. NiV was recovered from the urine of Pteropus bats in Malaysia, Cambodia, and Thailand (Iehle et al., 2007; Reynes et al., 2005; Wacharapluesadee et al., 2005). In Bangladesh, P. giganteus bats live in close proximity to human populations, often roosting in trees located in rural Bangladeshi villages. Thus, bat urine, intermittently laced with NiV, contaminates the immediate physical environment in many villages in Bangladesh. Yet in each of the 8 Nipah outbreaks investigated in Bangladesh, an association between living near a bat roost and infection with Nipah was looked for but was never found. This suggests that the quantity of viable virus shed in bat urine is too low or very few bats pass viable virus in urine to initiate clinically apparent infection in humans.
Eating bat-bitten fruit is often suggested as a pathway of transmission for human Nipah infection. NiV was recovered from fruit dropped by Pteropus bats in Malaysia (Chua et al., 2002). It is the most commonly suggested pathway for NiV transmission from bats to domestic animals. In contrast to general environmental contamination with urine, punctured fruit contaminated with bat saliva may favor virus survival. In Bangladesh where 43% of children under the age of 5 years meet the World Health Organization standards for chronic mal nutrition (NIPORT, 2007), little food is wasted. In outbreak investigations, villagers, especially children, commonly report consuming fruit which was partially eaten by bats. However, in the 6 NiV outbreak investigations where the question was asked, case patients never reported consuming partially eaten fruit significantly more than did controls.
____
Did outbreaks of human NiV infection occur in Bangladesh before the first outbreak was recognized in 2001?
Probably Yes.
Pteropus giganteus are widely distributed across Bangladesh (Nowak, 1994), and wherever Pteropus bats have been tested they have antibody to henipavirus (Epstein et al., 2008; Hsu et al., 2004; Iehle et al., 2007; Reynes et al., 2005; Wacharapluesadee et al., 2005). When Pteropus bats are experimentally infected with NiV they do not become clinically ill (Middleton et al., 2007), which suggests that NiV likely coevolved with its Pteropus hosts over millennia. Bangladesh has long been densely populated, and date palm sap harvesting is an old profession using techniques and simple tools that are passed on from father to son. Moreover, people frequently die in Bangladesh of unknown causes, often outside of hospitals. Three factors that have contributed to recognition of Nipah outbreaks recently include development of diagnostic tests for Nipah infection following the Malaysian outbreak, expansion of surveillance for a range of communicable disease by the government of Bangladesh, and expansion of news media coverage in rural Bangladesh.
Although NiV was discovered in Malaysia in 1999, retrospective investigations conducted later had suggested that NiV was responsible for sporadic disease in pigs in Peninsular Malaysia since late 1996, but was not recognized because the clinical signs were not markedly different from those of several endemic pig diseases, and because morbidity and mortality were not remarkable in pigs.
____
In Utero Transmission of Nipah Virus: Role played by Pregnancy and Vertical Transmission in Henipavirus Epidemiology, a 2007 study:
In utero transmission has been established for some viruses, including cytomegalovirus, varicella-zoster virus, rubella virus, poliovirus, Japanese encephalitis virus, coxsackie viruses, echovirus, measles virus, mumps virus, and hepatitis B, C, and E viruses. For other viruses, including HIV, delivery and postpartum transmission are the dominant pathways of vertical transmission. The importance of pregnancy and fetal materials in the spread of henipavirus disease was first hypothesized after the discovery that the index case of the 1994 HeV outbreak was a pregnant mare. This was later supported by the observation that HeV outbreaks have occurred during the birthing period in some species of flying foxes and by the isolation of virus from a pregnant flying fox and its fetus. Vertical transmission of HeV was later experimentally confirmed in guinea pigs and bats.
Mungall et al. report the results of a detailed investigation of in utero NiV transmission and provide the first experimental evidence that NiV, like HeV, can be vertically transmitted in cats. Cats are naturally infected and consistently exhibit characteristic disease pathology even at a low MOI. Multiplicity of infection (MOI) is a frequently used term in virology which refers to the number of virions that are added per cell during infection. If one million virions are added to one million cells, the MOI is one. A previous study has demonstrated the presence of HeV antigens both in adult tissues and in the placenta of infected pregnant flying foxes. However, despite rigorous sampling regimens, virus has been isolated only infrequently, and, where isolation was successful, positive sources have included urine and the fetus, heart, placenta, kidney, and spleen of 2 pregnant bats. Mungall et al. isolated significant amounts of infectious NiV from placental fluid (1 × 105 TCID50/mL) as well as from placental tissue, although at lower levels from the latter. Evidence was also provided for high levels of viral replication in many tissues of a pregnant adult cat and in fetal tissues, suggesting both vertical and horizontal transmission of this virus—a finding that has important implications for the epidemiology of NiV infection as well as for the testing of inhibitors and vaccines in this animal model and for the understanding of disease mechanisms. The pathology in cats resembled various features seen in humans, although encephalitis has not been observed in cats. Interestingly, temperature increase (as a measure of the development of infection) was initially the same for the pregnant cat and the infected nonpregnant control cat during the first 5 days; this was followed by a rapid increase for the control cat, whereas the infected pregnant cat showed a slight drop in temperature followed by a largely constant period until day 12. One can speculate that pregnancy delayed the progression to disease by 1 week, and perhaps the disease progression resumed as a result of the “secondary” infection in the fetus, in which the infection continued unchecked by maternal defense (immune) systems. If this is true, it represents a unique example of protection (although partial in this case) as a result of pregnancy. One can further speculate, as the Mungall et al. did, that such protection could be due to hormonal changes that occur during pregnancy. The data from this study were based only on 2 cats, and more experiments are required to confirm this novel observation. The results of Mungall et al.’s study have implications for the mechanisms of NiV spread in animal and human populations. The results affirm observations in horses, guinea pigs, and bats that demonstrate that henipaviruses can replicate to high titers in both adult and fetal tissues, indicating that both horizontal and vertical routes of transmission can play a role in spillover events. They also suggest a possible role for cats in HeV and NiV outbreaks that has never been fully investigated, even though cats were observed at the sites of both HeV and NiV outbreaks. Finally, the similarity in major pathological features between this cat model of disease and human infection could help to develop novel treatments, for example, by identification of maternal pregnancy factors that could delay progression to disease.
_____
_____
Spread of nipah:
The basic reproduction number, R0, is defined as the expected number of secondary cases produced by a single (typical) infection in a completely susceptible population. From past Nipah virus outbreaks, the R0 was estimated to be ~0.4. R0 is mathematical term quantifying the average number of new infections that one infected individual can generate, in an otherwise naïve population. For an infection to spread through a population, R0 needs to >1. When R0 is <1, the infection will eventually die out. Human-to-human transmission requires intimate contact with high levels of bodily secretions (respiratory secretions, saliva, urine, etc.). The risk of wide-spread transmission is therefore low. This is also reflected in estimated R0 ~0.4 for Nipah virus. Past Nipah outbreaks have been self-limiting because the virus doesn’t spread very easily from human to human. Outbreaks have tended to fizzle out after four or five chains of transmission in humans. But our worry each time there’s a new outbreak is that a mutation of Nipah virus may emerge that’s efficient at spreading among people.
The current Kerala outbreak appears to be small, and the appropriate public health measures have been rapidly implemented to contain its spread. To put the current Nipah virus outbreak in context, consider the following two vignettes:
- The 2001 outbreak in Siliguri, India, involved 66 people. The index case transmitted the virus to 11 additional patients at the hospital. These secondarily infected patients were transferred to other facilities – in two facilities, subsequent transmission involved 25 staff and 8 visitors. This was likely before the implementation of universal precautions— personal protective equipment (PPE) such as gloves, masks and/or face shields.
- 50% of Pteropus bats sampled in an outbreak area (Thakurgaon district) in Northwest Bangladesh were seropositive for Nipah virus antibodies. Yet transmission is still very sporadic. Thus, the drivers of virus spill-over remain relatively unknown (other than drinking of virus contaminated date palm sap).
_____
Is Nipah virus is as deadly as Ebola?
Yes, it’s comparable. Every outbreak has its own case fatality rate, and outbreaks can vary a lot in how easily they spread among people and how deadly they are, depending who gets infected and the genetic strain of the virus itself. But in general, Nipah tends to have an average case fatality rate of 75 percent—and some outbreaks have been 100 percent fatal. From that perspective, Ebola has had similar mortality rates.
Is Nipah the next Ebola?
No. While the mortality rate for those infected can be high, infection is not all that common. Before this latest outbreak, about 348 deaths had been linked to Nipah, most of which occurred in Southeast Asia and Bangladesh. In order for a disease to spread globally, each person has to infect at least more than one person. But a person with Nipah tends to infect either zero or one other person, according to a 2009 study published by the U.S. Centers for Disease Control and Prevention. By comparison, a person with measles can infect on average 10 others who aren’t vaccinated. And people who caught Ebola during the 2014 outbreak in West Africa tended to pass it on to between one and three others. But anytime the virus is inside a human, it has the opportunity to evolve and adapt to that human-specific environment. The worst-case scenario is a future strain that can transmit more quickly or easily among humans, which is why the WHO and global health experts are urging more research into vaccines and treatments.
______
Nipah virus worldwide spread:
The fact that the reservoir and intermediate hosts are Pteropus fruit bats and pigs, respectively, is a primary concern for researchers. Pteropus fruit bats are a migratory species capable of flying long distances. Pigs are also a primary export of Southeast Asia as well as a popular farm animal in the same region. The combination of these two key aspects of the Pteropus fruit bat and pigs makes the transmission of the Nipah virus to other parts of the world extremely easy. NiV infections have also been identified in cats, dogs, and horses.
_
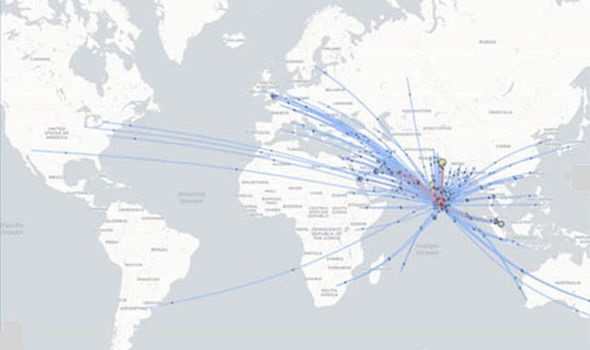
The map above shows countries likely to be hit by NiV.
EcoHealth Alliance, which created the above simulation, has warned that urgent action must be taken to prevent the outbreak turning into an epidemic. Theoretically it’s possible that, if Nipah cases make their way to a city where there is an international airport, someone infected with Nipah could get on airplane and go to another country. Kerala has airports in Calicut, Cochin, and Trivandrum, with Coimbatore International Airport in Tamil Nadu and Mangalore Airport in Karnataka also nearby – and all five offer international flights. Using FLIRT software developed by EcoHealth Alliance’s technology team, the global nonprofit organisation has undertaken a thorough analysis of which cities and countries should be on alert for potential influx of Nipah-infected travellers. The UK comes in 11th, the highest-ranked place outside Asia and ahead of China, in 14th, and the United States, in 17th. The simulation considered non-only direct flights, but also multi-leg journeys. Before long it has travelled to all six inhabited continents, if the virus spreads to any of the nearby states, as some fear it already has, the odds of Nipah becoming an international problem grow substantially. Goa, in particular, is a very popular vacation destination for many in Europe and Russia. Characteristics that might increase the risk of person-to-person transmission would be a virus that has a stronger tendency to move to the respiratory tract in high numbers. It is conceivable that the virus could acquire a mutation that would enhance this capacity. One concern is that any time a virus infects a human, it is in an environment that selects for survival in that context.
____
The pandemic potential of Nipah virus, a 2013 study:
- Nipah virus is a zoonotic paramyxovirus whose natural reservoir is bats of the genus Pteropus.
- Over half of people infected with Nipah virus die.
- Nipah virus is occasionally transmitted from person to person.
- Emergence of a Nipah virus strain with more efficient person-to-person transmission could produce a deadly global pandemic.
- High-income countries should support interventions to reduce person-to-person transmission in low-income countries.
Nipah virus, a paramyxovirus whose wildlife reservoir is Pteropus bats, was first discovered in a large outbreak of acute encephalitis in Malaysia in 1998 among persons who had contact with sick pigs. Apparently, one or more pigs was infected from bats, and the virus then spread efficiently from pig to pig, then from pigs to people. Nipah virus outbreaks have been recognized nearly every year in Bangladesh since 2001 and occasionally in neighboring India. Outbreaks in Bangladesh and India have been characterized by frequent person-to-person transmission and the death of over 70% of infected people. Characteristics of Nipah virus that increase its risk of becoming a global pandemic include: humans are already susceptible; many strains are capable of limited person-to-person transmission; as an RNA virus, it has an exceptionally high rate of mutation: and that if a human-adapted strain were to infect communities in South Asia, high population densities and global interconnectedness would rapidly spread the infection. Appropriate steps to estimate and manage this risk include studies to explore the molecular and genetic basis of respiratory transmission of henipaviruses, improved surveillance for human infections, support from high-income countries to reduce the risk of person-to-person transmission of infectious agents in low-income health care settings, and consideration of vaccination in communities at ongoing risk of exposure to the secretions and excretions of Pteropus bats.
_____
International norms and approaches in non-endemic countries:
Due to the significant morbidity and mortality, and rapid spread potential in domestic animals, and evidence of zoonotic properties, recently, Nipah virus has been included in the list of diseases with relevance for international trade of the World Organisation for Animal Health. The introduction of Nipah virus in non-endemic areas and in particular in Europe remains a plausible reality, primarily taking into account the presence of potentially NiV susceptible animal species. Intensive swine farming is widespread and transmission from pigs to humans was a key epidemiological feature of outbreaks in Malaysia and Singapore. Theoretically, wild boars might also play a role of amplification host. With concern to epidemiological perspectives, the presence in Europe of potentially NiV carrier bats of the species Daubenton’s bat (Mytotis daubentoni) represents another important aspect. Bat species in the genus Myotis naturally reside in trees, buildings, and caves that can be in close proximity to human residential areas, which increases the potential of transmission of zoonotic pathogens from bats to humans. Furthermore, susceptibility of ferrets to Nipah virus raises the possibility that the epidemiology could change further, evolving in natural condition and extending to other mustelids, and if a readily respiratory-transmissible Nipah virus could be created by serial passage in these wild animal species, as suggested by experiments in ferrets with H5N1 avian influenza. In summary, it cannot be excluded that the virus might be introduced and diffused through insectivorous bats, domestic pigs or other wild animals such as wild boars or mustelids, and finally might circulate in the human population on the base of person-to-person transmission capacity. Therefore, these elements suggest the importance to monitor the NiV epidemiological evolution, in terms of variation of geographical distribution and acquisition of new transmission ways (Table below).
_
Elements suggesting potential for NiV epidemiological changes with increasing impact on public health and animal health in currently free countries, thus justifying monitoring of epidemiological evolution:
| Possible diffusion of Nipah Virus in free countries? |
| Fruit bats absent – Insectivorous bats? |
| Transmission from pig to man – Wild boar? |
| Ferrets sensible to NiV – Other mustelids? |
| Person-to-person direct transmission |
_
In conclusion, knowledge and awareness on the disease should be improved and disseminated to health services, veterinarians, farmers and consumers. Nipah virus, as other zoonotic agents, might be included in monitoring plans, in particular for wild animals. Prioritization may drive the attention to other pathogens showing for example higher incidence in the population. However, field investigations may demonstrate radical and unexpected epidemiological changes. For example, the discovery of a novel ebolavirus-like filovirus in Spanish microbats demonstrated that the potential for such spill over events is not limited to Africa or Asia. It is therefore important to enhance our preparedness to counter potential future introduction of exotic pathogens as Henipaviruses in non-endemic areas by conducting active pre-emergence research. Of utmost importance, monitoring the evolving epidemiology of a dangerous pathogen like the Nipah virus is an essential element to be able to promptly adapt control plans in the case that it might become a new public health priority.
______
______
Nipah incubation period and infectivity:
The incubation period for both Nipah and Hendra is different among infected humans and animals. In animals, the incubation period is 7–14 days for Nipah and 5–16 days for Hendra after exposure. Among humans, the incubation periods are 4–20 days and 5–12 days, for Nipah and Hendra, respectively. Although average incubation period for nipah is 4 to 20 days in humans; it can be as short as two days or as long as 60 days. 40% to 100% of clinically apparent cases die. There are many clinically unapparent (sub-clinical) cases. Some people with mild or subclinical infections can develop late-onset encephalitis months or years later. Mortality in pigs is only 5% but transmission is 100%.
Infectious dose: unknown.
Initiation of infection by a single poliovirus particle depends on translation of the +RNA, self-cleavage of the polyprotein, processing to functional polioviral enzymes, and amplification of the RNA. The poliovirus +RNA is handicapped initially; a single +RNA must compete with millions of cellular mRNA’s for translation. Although a single polioviral +RNA might be sufficient to initiate an infectious cycle, the probability of it being translated in the presence of millions of mRNA’s with a cap advantage is very low. Boost the number of poliovirus +RNA’s in the cell a thousand-fold and the probability of successful translation is increased (a thousand-fold). Once that happens, it is all downhill for the cell and uphill for poliovirus! The poliovirus protease knocks out the capped mRNA advantage and polio RNA takes over the cell machinery. This is the concept of infectious dose. Infectious dose of influenza A is approximately 790 organisms via the nasopharyngeal route. Infectious dose of Ebola is 10 organisms by contact transmission. A study found that the minimal NiV infectious dose to infect ferrets oronasally is 5×103 TCID50 with highest virus load in the lungs and spleen, detected in all animals infected with this and one log10 higher dose. 50% Tissue culture Infective Dose (TCID50) is the measure of infectious virus titer. This endpoint dilution assay quantifies the amount of virus required to kill 50% of infected hosts or to produce a cytopathic effect in 50% of inoculated tissue culture cells. The minimal infectious dose by aerosol inoculation is 0.6–3.0 TCID50 for influenza A in humans.
Route of entry: A portal (route) of entry is the site through which micro-organisms enter the susceptible host and cause disease/infection. Infectious agents enter the body through various portals, including the mucous membranes, the skin, the respiratory and the gastrointestinal tracts. Respiratory tract is the main route of entry of NiV in humans via inhalation of infected droplets using nasopharyngeal & oropharyngeal mucosa and small airways. Oropharyngeal mucosa is also used for entry of NiV from contaminated date palm sap, contaminated fruits etc. when ingested. Virus enters brain via blood and/or via olfactory nerve. Accidental needle prick injury apparently did not transmit NiV but close contact with infected patient did transmit disease proving that skin and mucous membrane could also be route of entry using secretions and excretions of infected patient.
Contagiousness: Nipah virus is moderately contagious as it needs close contact and usually infects the family members and/or medical caregivers of NiV-infected individuals. The estimated R0 is ~0.4 for Nipah virus. Compared with other diseases such as tuberculosis, smallpox, measles, chicken pox, influenza, SARS and polio, nipah is much less contagious.
Period of communicability of Nipah virus infection: Unknown. Doctors do not completely understand the contagious period for infection, but it likely begins during the incubation period (proven in pigs) and continues until the patient stops shedding virus. In most patients, this occurs when the symptoms and signs of the infection are diminished or gone. The infected patient is maximally contagious when patient is gravely ill.
Susceptibility and resistance to Nipah virus infection: Undetermined. Relapses of infection can occur.
Mode of transmission (vide supra): The mechanism for the transmission of the virus from fruit-bats to animals is unknown, but may involve consumption of fruit contaminated with urine or saliva from infected bats. Transmission from animals to humans appears to occur by direct contact with contaminated tissues/body fluids of infected animals, especially pigs. Human to human transmission is likely to occur by direct exposure to an infectious inoculum shed in the respiratory secretions of the infected individual, as well as by close physical interaction and frequent contact with the infected individual’s saliva.
_
Asymptomatic nipah infection in humans:
Nipah virus can cause asymptomatic infections in some patients; in the Malaysia and Singapore outbreaks between 17% and 45% of infections were asymptomatic. There has not been any evidence for asymptomatic infection in outbreaks in Bangladesh although cases of mild illness were identified. (Hsu et al., 2004). In a study of Malaysian households with symptomatic family members infected by Nipah virus, 6 of the 36 (17%) antibody positive individuals were asymptomatic (Tan et al., 1999). Parashar found that among symptomatic pig farmers and their families in Malaysia, 30 of 110 (27%) were asymptomatic (Parashar et al., 2000). In another study designed specifically to compare symptomatic versus asymptomatic Nipah virus infection among high risk groups in Singapore, 10 of 22 (45%) of those with Nipah virus antibodies were asymptomatic, with no neurologic or respiratory symptoms (Chan et al., 2002).
_____
_____
Clinical presentation of Nipah virus disease:
In humans, NiV causes encephalitis (inflammation of the brain) and/or respiratory signs and symptoms. The case-fatality rate of encephalitis is estimated at 40–75 per cent. Human cases range from asymptomatic to cases involving fatal encephalitis. Infected people initially develop a flu-like illness with fever, headaches, myalgia (muscle pain), vomiting and sore throat. This can be followed by dizziness, drowsiness, altered consciousness, and neurological signs of acute encephalitis. Encephalitis and seizures occur in severe cases, progressing to coma within 24–48 hours. Some people can develop atypical pneumonia and severe respiratory problems, including acute respiratory distress syndrome. Most people who survive acute encephalitis make a full recovery, but approximately 20 per cent are left with residual neurological deficits, such as persistent convulsions and personality changes. A small number of people, usually those with mild or subclinical cases, can relapse or develop delayed-onset encephalitis.
_
Signs and symptoms observed in these patients in order of frequency are: fever (100%), altered mental status (90%), headache (73%), weakness (67%), respiratory distress (69%), cough (62%), diarrhea (29%), seizures (23%). You should be ruled out this infection in people from the regions of Southeast Asia where nipah outbreaks have occurred.
Clinical features of nipah:
Symptoms:
The following symptoms were observed (in order of frequency in Bangladeshi cases)
- Fever
- Altered mental status
- Severe weakness
- Headache
- Respiratory distress
- Cough
- Vomiting
- Muscle pain
- Convulsion
- Diarrhoea
General Signs:
- Reduced GCS score
- Raised temperature
- Increased respiratory rate (Adult: ≥25/min; children of ≥ 12 months: ≥ 40/min)
- Increased heart rate (Adult: ≥100/min; children of ≥ 12 months: ≥ 140/min)
- Crepitations in lung
- Hypertension/Hypotension
Neurological signs:
-Oculoparesis
-Pupillary abnormality
-Facial weakness
-Bulbar weakness
-Limb weakness
-Reduced deep tendon reflexes
-Plantar-absent/extensor
___
Malaysian authorities studied 94 patients with Nipah virus infection, the majority (93%) of whom had direct contact with pigs 2 weeks before the onset of illness. This implied a short incubation period. Fever, headache, dizziness, and vomiting featured prominently. Reduced levels of consciousness and brainstem dysfunction were present in 52 patients (55%). Segmental myoclonus, areflexia, hypotonia, hypertension, and tachycardia were other features. Antibodies against Nipah virus were detected in serum or cerebrospinal fluid (CSF) in 76% of 83 patients tested. Severe brain-stem involvement resulted in the deaths of 30 patients (32%). Fifty patients (53%) achieved full recovery, while neurological deficits persisted in 14 (15%). Three patients relapsed neurologically after initial mild disease, a finding that is unique to Nipah encephalitis. Apart from the CSF lymphocytic predominance, other laboratory markers were not specific. These included abnormalities like moderate thrombocytopenia and elevation of liver transaminases.
The cases in Singapore manifested similarly except for the finding of abnormal chest radiographs in 8 of the 11 patients. In two of these individuals there was evidence of an atypical pneumonia. Serological tests for mycoplasma, chlamydia, Q fever, and legionella were negative on acute and convalescent sera. Curiously, one of these patients with atypical pneumonia developed a transient episode of visual hallucinations during which he described seeing pigs flying outside his hospital room! One of the three symptomatic patients discovered during the screening of abattoir workers had mild neurological deficit. It turned out that he had been hospitalized for an atypical pneumonia coincidentally during the outbreak. Thus, this disease may present initially as pneumonia without any neurological feature.
____
Clinical Presentation of Nipah Virus Infection in Bangladesh, a 2008 study:
In Bangladesh, 7 outbreaks of Nipah virus infection were identified during the period 2001–2007. In Bangladesh, Nipah virus infection was associated with contact with a sick cow, consumption of fresh date palm sap (potentially contaminated with pteropid bat saliva), and person-to-person transmission. The Malaysian outbreak was associated with a single strain. By contrast, viruses isolated in Bangladesh represent diverse strains. Hypothetically, a single strain of Nipah virus could result in a narrower range of clinical presentations than those found during epidemics associated with genetically diverse strains. Thus, Nipah virus illnesses occurring in Bangladesh potentially provide insight into broader clinical manifestations of Nipah virus infection. Authors describe the clinical presentation of 92 Nipah virus-infected patients identified during the first 4 outbreaks in Bangladesh during the period 2001–2004. Authors identified 92 patients with Nipah virus infection, 67 (73%) of whom died. Although all age groups were affected, 2 outbreaks principally affected young persons (median age, 12 years); 62% of the affected persons were male. Fever, altered mental status, headache, cough, respiratory difficulty, vomiting, and convulsions were the most common signs and symptoms; clinical and radiographic features of acute respiratory distress syndrome of Nipah illness were identified during the fourth outbreak. Among those who died, death occurred a median of 6 days (range, 2–36 days) after the onset of illness. Patients who died were more likely than survivors to have a temperature >37.8 DegC, altered mental status, difficulty breathing, and abnormal plantar reflexes. Among patients with Nipah virus infection who had well-defined exposure to another patient infected with Nipah virus, the median incubation period was 9 days (range, 6–11 days). Nipah virus infection produced rapidly progressive severe illness affecting the central nervous and respiratory systems. Clinical characteristics of Nipah virus infection in Bangladesh, including a severe respiratory component, appear distinct from clinical characteristics reported during earlier outbreaks in other countries.
____
Factors associated with the Prognosis of Nipah Virus Infection.
Factors that appeared to affect the survival of patients are summarized in Table below.
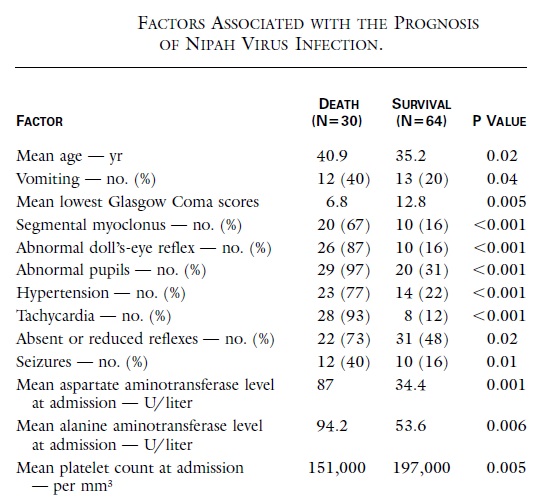
As compared with the patients who survived, the patients who died were older and had more severe brain-stem involvement, as evidenced by the presence of a reduced level of consciousness, vomiting, abnormal doll’s-eye reflex, abnormal pupils, hypertension, and tachycardia during the course of the illness. Other poor prognostic factors were the presence of segmental myoclonus, seizures, areflexia, more greatly elevated hepatic aminotransferase levels at admission, and lower platelet counts at admission. There appeared to be no significant difference in outcome with ribavirin treatment. Multivariate logistic-regression analysis showed that an abnormal doll’s-eye reflex (P=0.025) and tachycardia (P<0.001) were significantly associated with mortality. The estimated probability of mortality in the presence of both factors was 0.84, whereas it was 0.02 in the absence of both factors.
______
______
Pathogenesis and pathology of nipah virus disease:
The natural reservoir hosts of Hendra virus and Nipah virus are several species of pteropid fruit bats among which they are not known to cause disease (Halpin et al., 2011). However, Hendra and Nipah viruses possess an exceptionally broad species tropism and both natural and experimental infections have demonstrated their capacity to cause disease which can often be fatal in horses, pigs, cats, dogs, ferrets, hamsters, guinea pigs, monkeys, and humans, spanning 6 mammalian Orders. In disease susceptible animal hosts and people, Nipah virus and Hendra virus cause a systemic infection that is characterized as a wide-spread vasculitis and endothelial cell tropism. Though this pathology is not unique to these henipaviruses, an understanding of Hendra and Nipah virus cellular tropism on the molecular level has provided an explanation to this disease feature which includes the appearance of syncytia, thrombosis, ischemia and necrosis, with parenchymal cell infection and associated pathology in many major organ systems, and prominently in the brain and lung. The major involvement of the lung and brain in Hendra and Nipah virus infection often manifests as an acute severe respiratory syndrome, encephalitis or a combination of both. Disturbingly however, infection in people can also have longer term consequences, and in addition to an acute symptomatic infection, Hendra and Nipah virus infection can also take a protracted course following recovery from an initial infection. Individuals in these cases can later undergo a recrudescence of virus replication in the central nervous system (CNS) causing a relapse of encephalitis, a process that was first noted in the second fatal case of Hendra virus human infection (O’Sullivan et al., 1997; Wong et al., 2009). Quite remarkably, relapsed-encephalitis caused by Nipah virus has been reported in people from several months to as long as 11 years following infection (Abdullah et al., 2012). How the henipaviruses survive immune-mediated clearance and can later cause a recrudescence of replication in the CNS is unknown, but this virological feature clearly has important implications for anti-henipavirus therapeutics development.
_
The site of primary replication of NiV and the mode of virus propagation throughout the organism remain unknown. The incubation period varies from 4 to 60 days and is shorter in Bangladeshi than in Malaysian patients, potentially reflecting differences between two viral strains. In humans, the blood vessels appear to be one of the early targets of infection, with the central nervous system (CNS) being the most severely affected, although lung, kidney, and other organs are also infected. The majority of human infections led to acute encephalitis with vasculitis-induced thrombosis in the brain and, in some patients, atypical pneumonia and respiratory distress. A late-onset encephalitis could arise up to several months or even years after the initial infection, and a relapsed encephalitis has occurred in patients who had previously recovered from acute encephalitis.
_
Pathology played a key role in identifying the causative agent. Histopathologic findings include vasculitis with thrombosis, microinfarctions, syncytial giant cells, and viral inclusions. Although characteristic of this disease, syncytial giant endothelial cells are seen only in 25% of cases, and viral inclusions of similar morphology can been seen in other paramyxoviral infections. Immunostaining provides a useful tool for unequivocal diagnosis of the disease, demonstrating viral antigen within neurons and endothelial cells of most organs.
_
Samples collected at autopsy from patients with positive results of cultures for Nipah virus showed histologic findings of endothelial damage and vasculitis, mainly in arterioles, capillaries, and venules. Autopsy studies revealed the central pathology of this disease to be vasculitis-induced thrombosis causing ischemia and infarction. Multi-organ necrotizing vasculitis and syncytial formation were characteristic. This was especially so in the central nervous system where a diffuse small blood vessel vasculitis and lytic necrosis were evident in the cerebral cortex and brainstem associated with infection of endothelial cells. The brain was the most severely affected organ, but other organs, including the lungs, the heart, and the kidneys, were also affected. Vasculitis of blood vessels was characterized by vessel-wall necrosis, thrombosis, and inflammatory cell infiltration of neutrophils and mononuclear cells. Syncytial cell formation was seen in the endothelium of affected blood vessels in the brain and lungs and in the Bowman’s capsule of the glomerulus. Zones of microinfarction and ischemia were commonly found surrounding or adjacent to vasculitic blood vessels. In the brain, many neurons had eosinophilic cytoplasmic and nuclear viral inclusions, as is seen in association with other paramyxovirus infections.
_
Mechanism of Pathogenesis of Nipah virus infection:
Widespread vasculitis, a key event in the pathogenesis of Nipah virus infection, seems to be a consequence of infection of the vascular endothelial and smooth muscle cells. Overall, the frequency of vasculitis seemed to be proportional to necrosis and necrotic plaques, particularly in the CNS and lung. The necrotic plaques and the acute encephalitic syndrome may stem from both direct neuronal infection and ischemic injury as seen in the figure below. This sequence of pathological events is supported by the concomitant increase in frequency of syncytia, vasculitis, thrombosis, necrotic plaques, and viral antigen in the CNS.
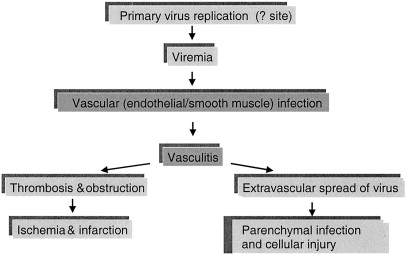
The pattern of vasculitis and viral antigen distribution suggests that endothelial infection occurs before transmural spread to underlying smooth muscle in the tunica media. Finding of numerous infected neurons in association with necrotic plaques and vasculitic vessels suggests that a breach of the blood-brain-barrier may facilitate virus escape and extravascular CNS infection. In subacute sclerosing panencephalitis caused by measles virus, endothelial infection is also thought to facilitate initial virus entry into the brain. Once neurons are infected, interneuronal spread along neuronal pathways may contribute to viral dissemination. Viral spread through damaged vessels may also explain non-CNS parenchymal infection.
__
The current understanding of routes of infection and virus spread throughout the host infected with henipaviruses is briefly summarized in figure below. Despite some differences in the pathogenesis and pathology between species, there are common characteristics of henipavirus infections, such as the development of viremia; the involvement of the respiratory, central nervous, and immune systems; vasculitis; the formation of syncytia in the endothelium; the infection of dendritic cells, etc. Considering the routes of infection, mucosal immunity may be important early postinfection; however, this has not been widely explored, even in terms of vaccine design.
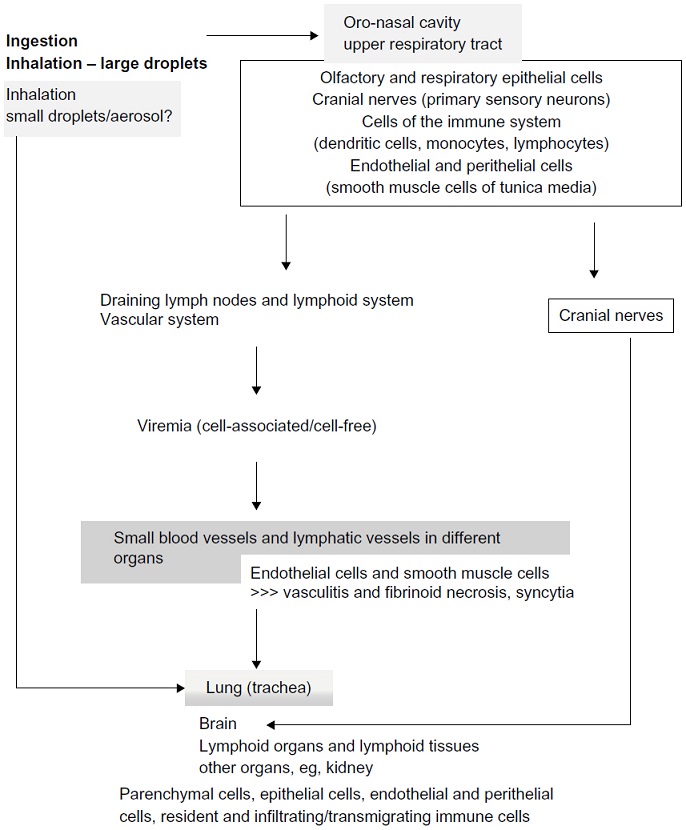
Figure above shows generalized summary of the current understanding of pathogenesis. The figure represents an overall picture covering a number of susceptible species.
__
Molecular mechanisms of NiV pathogenesis:
During the late stages of disease, virus replication spreads from the respiratory epithelium to the endothelium in the lungs. The infection can sometimes trigger a prominent vasculitis in small vessels and capillaries as characterized by endothelial syncytium and mural necrosis. Large vessels are usually not affected. NiV can then enter the bloodstream and disseminate throughout the host in either free form or by binding host leukocytes. In addition to the lungs, important target organs are the brain, spleen and kidneys, and viremia following respiratory infection can lead to multi-organ failure. Interestingly, NiV has been shown to bind to CD3+ leukocytes without entry or replication of the virus. In hamsters, passive transfer of NiV loaded leukocytes to naïve animals results in lethal infection. In contrast in pigs, NiV can productively infect monocytes, CD6+CD8+ T lymphocytes and NK cells. CD6 is a ligand for the activated leukocyte cell adhesion molecule ALCAM (CD166), which is highly expressed on the microvascular endothelial cells of the blood-air and the blood-brain barrier. NiV infection of T cells expressing CD6 may explain the preferential tropism of NiV for small blood vessels of the lung and brain. It is currently unknown whether binding of NiV or infection of human leukocytes will affect the phenotype of the cells, such as increase CD6 expression, thereby preferentially homing to the CNS.
Entry into the CNS is thought to occur through two distinct pathways: antegradely via the olfactory nerve and/or via the haematogenous route through the choroid plexus and cerebral blood vessels. Infection of the CNS in humans is characterized by vasculitis, thrombosis, parenchymal necrosis, and presence of viral inclusion bodies. Plaques with necrosis are found in both the gray and white matter and vasculitis, thrombosis, and parenchymal edema and inflammation are found in the vicinity of these plaques. Inflammatory cells found in the CNS primarily consist of neutrophils, macrophages, lymphocytes, and reactive microglia. NiV antigen can typically be detected in neurons and neuronal processes and endothelial cells. Occasionally, viral antigen is also detected in ependymal cells and rare glial cells in the white matter.
NiV infection of the CNS and the development of neurological signs are associated with the disruption of the blood-brain barrier (BBB) and expression of TNF-α and IL-1β. These pro-inflammatory cytokines have been shown to play a role in increasing the permeability of the blood-brain barrier as well as the induction of neuronal injury and death. While the source of TNF-α and IL-1β expression in the brain is currently unknown, they can be released by microglia, which are also infected by NiV. However, whether disruption of the BBB is a direct cytopathic effect of virus replication in the microvasculature or an indirect effect through expression of TNF-α and IL-1β by bystander cells such as neurons and microglia remains unclear.
Experimental studies in various animal models have shown direct entry of the CNS by NiV, through the olfactory nerve. In these models, NiV infects the olfactory epithelium in the nasal turbinates. NiV subsequently infects neurons extending through the cribriform plate into the olfactory bulb, providing a direct route entry into the CNS. NiV then disseminates to the olfactory tubercle and throughout the ventral cortex. It is currently unknown whether this route is also biologically relevant in human infections, since the olfactory epithelial surface is relatively large in these species compared to man.
_____
Comparison of the Pathogenicity of Nipah Virus Isolates from Bangladesh and Malaysia in the Syrian Hamster, a 2013 study:
Authors compared NiV-M and NiV-B infection and disease progression using the Syrian hamster model. They found that NiV-M is more destructive in cultured hamster cells and has faster onset of cytopathogenicity compared to NiV-B. This is also true in hamsters, where although both viruses are pathogenic and cause a similar disease, pathology caused by NiV-M infection is accelerated. These data show that there is a difference in disease progression between the two strains of Nipah virus and will allow for a more detailed understanding of the events leading to disease caused by these viruses.
_______
The immunomodulating V and W proteins of Nipah virus determine disease course, a 2016 study:
The viral determinants that contribute to Nipah virus (NiV)-mediated disease are poorly understood compared with other paramyxoviruses. Here authors use recombinant NiVs (rNiVs) to examine the contributions of the NiV V and W proteins to NiV pathogenesis in a ferret model. They show that a V-deficient rNiV is susceptible to the innate immune response in vitro and behaves as a replicating non-lethal virus in vivo. Remarkably, rNiV lacking W expression results in a delayed and altered disease course with decreased respiratory disease and increased terminal neurological disease associated with altered in vitro inflammatory cytokine production. This study confirms the V protein as the major determinant of pathogenesis, also being the first in vivo study to show that the W protein modulates the inflammatory host immune response in a manner that determines the disease course.
______
______
Nipah virus and immune system:
Researchers have analyzed the permissiveness of human leukocytes to NiV infection. Only dendritic cells (DC) support a productive NiV infection, although all leukocyte types are capable of binding NiV and transmitting it to susceptible cells in vitro. Human lymphocytes and monocytes are not permissive for NiV and a low level of virus replication is detected only in dendritic cells. Interestingly, despite the absence of infection, lymphocytes could efficiently bind NiV and transfer infection to endothelial and Vero cells. This is called lymphocyte-mediated transinfection. Although NiV binds to lymphocytes, it is not internalized, since it can be stripped off by proteolysis and remains sensitive to neutralization by antibodies. The virus stays bound at the cell surface for several days without much loss of infectivity compared to cell-free virus, as if stabilization occurs upon contact with the plasma membrane. Upon contact, cell-bound virus is quickly transferred to a permissive cell. Although monocytes and macrophages attach NiV, their lower level of transinfection may be associated with the high phagocytosis capacity of these cell types, leading to the decreased cell surface exposure of viral particles and their likely inactivation upon endocytosis. Accordingly, in the hamster infection model, virus is found associated with leukocytes without signs of infection, and the transfer of these cells resulted in the infection of naïve animals, demonstrating efficient virus transinfection in vivo. NiV may initially enter the respiratory or digestive tract, potentially via abrasions and breaks in the mucosal surface or skin, where it could infect local dendritic cell (DC). Infected DCs migrate to regional lymph nodes via the lymphatic vessels, as described for measles, another paramyxovirus. There, locally produced virus binds to lymphocytes, which subsequently exit the lymph nodes and act as passive vehicles for NiV spread to susceptible cells of the blood vessels of many target organs and possibly enable NiV to cross the blood brain barrier to cause fatal encephalitis. NiV binding to lymphocytes may provide protection to virions and eventually allow their more efficient transport to different tissues, thus playing a critical role in NiV pathogenesis. The leukocyte-bound virus represents a potential target for therapeutic intervention, particularly in the early stages of infection. Indeed, the efficiency of anti-NiV antibodies has been demonstrated in hamsters and ferrets. Neutralizing antibody titers appear 7–10 days post-experimental infection with maximum titers seen 14–16 days post-infection. Ferrets administered a HeV soluble G protein vaccine were protected against live NiV challenge 14 months post-vaccination.
_
The rapid course of NiV infection, affecting multiple organs, indicates dissemination of the virus via the bloodstream. The present observations argue in favor of leukocytes acting as virus carriers, mediating transinfection of permissive host target cells. In humans, NiV has been isolated from urine and cerebrospinal fluid from infected patients, but there is no report of isolation from serum. Thus, peripheral blood mononuclear cells appear much more appropriate than serum for NiV diagnosis and virus isolation from infected patients. The lymphocyte-bound NiV particles are remarkably stable and retain infectivity for a prolonged period. The ability of leukocytes to transfer virus from infected to noninfected hamsters argues for the in vivo relevance of cell-mediated transinfection for virus spread throughout the organism. The high trafficking capability of lymphocytes and their constant interaction by rolling onto the endothelial cells could explain disseminated microvascular NiV infection in multiple organs in the absence of detectable viremia. This method of virus propagation may not be unique to NiV but could be used for dissemination of other viruses, such as the closely related Hendra virus, contributing to their high pathogenicity.
_
The IFN response through IFN-α and -β plays a critical role in controlling viral infections by signalling both infected and non-infected cells to enter into an antiviral state. This signalling occurs through the Janus Kinase (JAK)/Signal Transducer and Activator of Transcription (STAT) pathway. Many viruses have evolved means of inhibiting IFN signalling. Paramyxoviruses have evolved mechanisms whereby the P gene products (P, V, W and C) inhibit the JAK/STAT signalling pathway through a common N-terminal domain that can bind to STAT1 and inhibit its phosphorylation. Non-structural proteins of a large number of viruses are known to have inhibitory effects on IFN signalling. In the case of NiV, V proteins have been demonstrated to subvert IFN responses by sequestering STAT1 and STAT2 in high molecular mass cytoplasmic complexes without inducing degradation and the complex formation prevents IFN-induced STAT tyrosine phosphorylation. The N terminal domain of the V protein, from residues 100 to 160, is responsible for the activity and the region is shared by the W and P proteins. Furthermore, the single amino acid at residue 125 plays a critical role in IFN antagonism exerted by NiV V protein. The NiV W protein has a nuclear localization signal in its unique terminal domain and sequesters STAT1 in the nucleus, thus creating both a cytoplasmic and a nuclear block for STAT1, and it inhibits the transcription of antiviral genes under control of ISRE promoters activated by ISGF3. Thus, P, V and W proteins act as antagonists for IFN-responses by sequestering STAT proteins, with the W protein being the most efficient and the P protein the least. NiV V and W proteins can also inhibit virus-induced activation of the IFN-β promoter and IRF3-responsive promoter (ISG54 promoter) and only the W protein shows strong inhibition of promoter activation in response to the stimulation of Toll-like receptor 3. NiV C protein was also suggested to have weak IFN antagonist activity, but the target is unclear. These antagonistic activities of IFN-signaling of P, V, W and C of NiV were all demonstrated by the experiments using transiently expressed proteins following transfection with plasmid DNA, and researchers also confirmed strong inhibition of IFN response with P, V, W and C proteins expressed by plasmids in the reporter gene assay. A study shows that that NiV-C shuttles between the nucleus and cytoplasm. Authors also clarified that NiV-C has nuclear export signal and nuclear localization signal using NiV-C deleted, alanine substitution mutants and enhanced green fluorescent protein (EGFP) fused proteins. And they also showed that interferon (IFN) antagonist activity of NiV-C related to its subcellular localization. The results indicate that NiV-C exert IFN antagonist activity in the cytoplasm.
_
NiV and Complement system:
The complement system constitutes a complex group of soluble and cell-associated proteins that together form an integral part of the innate host defense against pathogens. Complement serves to link innate and adaptive immunity to viruses through recognition of virions, direct neutralization of infectivity, recruitment and stimulation of leukocytes, opsonization by immune cells, and activation of T and B cell responses. Complement activation plays important roles in viral pathogenesis and has been the focus of efforts to improve the effectiveness of vaccines and therapeutic vectors. The complement cascade can be initiated through three main pathways: the classical pathway, lectin pathway, or alternative pathway. These three pathways converge on a central component, C3, which is activated by cleavage into the anaphylatoxin C3a and into C3b, which can bind covalently to viral components to aid in opsonization and phagocytosis. Under normal conditions, inappropriate complement activation is regulated by a complex series of host proteins, with one key regulatory step being at the formation and stability of the C3 convertases. The key C3 convertase complex is also a common target for inhibition by many pathogenic microbes. For example, some enveloped viruses recruit host cell membrane-bound regulators into their envelope (e.g., CD55), which can then act to dissociate C3 convertase complexes that form on the virion surface. Viruses can also block the C3 convertase by exploiting factor I protease to cleave C3b or C4b into the inactive forms. Complement is strongly activated by a wide range of negative-strand RNA viruses, including parainfluenza virus 5 (PIV5), mumps virus (MuV), vesicular stomatitis virus (VSV), and Newcastle disease virus (NDV). For each of these viruses, C3 has been shown to play an essential role in effective in vitro neutralization. Conversely, many of these negative-strand RNA viruses exploit normal host cell inhibitors to evade complement pathways. NiV infection of humans is systemic, where viral antigen can be detected in a number of organs, including the respiratory tract, neurons, lymphoid tissue, and endothelial cells lining blood vessels. NiV can disseminate through binding to CD3+ lymphocytes. Because complement factors can be derived from serum, cells lining the respiratory tract, or by recruited lymphocytes, the sites of NiV infection and mechanisms of dissemination suggest that complement is a highly relevant innate immune pathway that NiV encounters during natural infections. NiV utilizes a virion-associated factor I-like protease to evade human complement pathways.
_____
_____
Case Detection, Investigations and Diagnosis of nipah virus disease:
_
Nipah case definition:

__
Suspected case:
A person fulfilling both of the following criteria is defined as a suspected case:
- Features of acute encephalitis as demonstrated by
-Acute onset of fever and
-Evidence of acute brain dysfunction as manifested by
Altered mental status OR
New onset of seizure OR
Any other neurological deficit
- Epidemiological linkage
-Drinking raw date palm sap OR
-Occurring during Nipah season OR
-Patient from Nipah endemic area OR Nipah outbreak area
Probable case:
A person with features of acute encephalitis
- during a Nipah outbreak in the area OR
- with history of contact with confirmed Nipah patient
In both suspected and probable cases, the patient might present with respiratory features with or without encephalitis. The respiratory features are
- Illness < 7 days duration and
- Acute onset of fever and
- Severe shortness of breath, cough and
- Chest radiograph showing diffuse infiltrates
Confirmed case:
A suspected or probable case with laboratory confirmation of Nipah virus infection either by:
- IgM antibody against Nipah virus by ELISA in serum or cerebrospinal fluid
- Nipah virus RNA identified by RT-PCR from respiratory secretions, urine, or cerebrospinal fluid
_
Definition of Cluster:
Two or more suspect cases living within a 30 minute walk of each other who develop symptoms within 21 days of each other.
_
Definition of a Contact:
A close contact is defined as a patient or a person who came in contact with a Nipah case (confirmed or probable cases) in at least one of the following ways.
- Was admitted simultaneously in a hospital ward/ shared room with a suspect/confirmed case of Nipah
- Has had direct close physical contact with the suspect/confirmed case of Nipah during the illness including during transportation.
- Has had direct close contact with the (deceased) suspect/confirmed case of Nipah at a funeral or during burial preparation rituals
- has touched the blood or body fluids (saliva, urine, vomitus etc.) of a suspect/confirmed case of Nipah during their illness
- has touched the clothes or linens of a suspect/confirmed case of Nipah.
These contacts need to be followed up for appearance of symptoms of NiV for the longest incubation period (21 days).
_____
Laboratory Investigations:
Thrombocytopenia (defined as a platelet count of less than 140,000 per microliter) was found in 30 percent of patients and leukopenia (defined as a white-cell count of less than 4000 per microliter) in 11 percent. Blood urea, creatinine, and electrolyte levels were normal in all patients. Elevated levels of alanine aminotransferase (>65 U per liter) were found in 33 percent of patients, and elevated levels of aspartate aminotransferase (>37 U per liter) were found in 42 percent of patients.
Cerebrospinal fluid examination was carried out in 92 patients in one study. The results of initial and repeated examinations were abnormal (as indicated by elevated white-cell counts, elevated protein levels, or both) in 75 percent and 77 percent, respectively. There was no correlation between abnormal cerebrospinal fluid findings and the severity of disease. The mean Glasgow Coma Scale score at nadir was 11.3 in those with abnormal findings and 9.5 in those with normal findings (P=0.1).
______
______
Laboratory diagnosis of Nipah:
In the diagnosis of nipah infection is necessary to rule out other infections that can be detected in patients from countries found infection with this virus, such as malaria, Japanese encephalitis, West Nile virus, Dengue infection, Hantavirus, or Leptospira infection.
_
Nipah virus infection can be diagnosed by a number of different tests. Since Nipah is classified as a biosafety level 4 (BSL4) agent, special precautions must be undertaken in the collection, submission and processing of samples. Biosafety considerations require that this work be carried out only in a physical containment level 4 (PC4) facilities. Various strategies have been developed to reduce the risk of laboratory sera, including gamma-irradiation or sera dilution and heat-inactivation. Henipavirus antigens derived from tissue culture for use in ELISA can be irradiated with 6 kilo Greys prior to use, with negligible effect on antigen titre.
_
Only a few laboratories worldwide have the capability for testing and confirming the presence of Hendra and Nipah viruses by virus isolation, immunohistochemistry, and molecular amplification. In addition, these viruses are classified as biosafety level 4 (BSL-4) agents and must be handled under strictest physical containment standards.
Viral isolation in cell culture from affected tissue is an important diagnostic methodology for these viruses, particularly when determining the etiology of a new outbreak (Daniels et al., 2001). Both Hendra and Nipah viruses grow well in Vero cells, and a cytopathic effect is usually noted within 3 days. Nipah virus has been isolated from human CSF, nasal and throat swabs, and urine (Chua et al., 1999; Goh et al., 2000). The virus has also been isolated from pigs and cats in a variety of tissue including lung, spleen, serum, and kidneys (Daniels et al., 2001; Middleton et al., 2002). Although Hendra virus has been isolated in humans only from kidney tissue, it has been isolated from serum, lung, spleen, and CNS tissue from a variety of animals including horses, cats, and guinea pigs (Murray et al., 1995a; Williamson et al., 1998, 2000, 2001). Immunostaining, neutralization techniques, PCR, and EM, including immunoelectron microscopy, are utilized for further identification of the virus.
_

_
Identification of the agent:
Virus isolation should be performed for definitive diagnosis in an area with a newly suspected outbreak. Experimentally, NiV is detectable in oropharyngeal and nasal swabs as early as two days post-infection; experimentally infected animals continued to shed virus until three weeks post-infection. Virus isolation by cell culture can be performed from brain, lung, kidney and spleen samples transported at 4°C in 48 hours or frozen if over 48 hours, using African green monkey kidney (Vero) and rabbit kidney (RK-13) cells. Cytopathic effect (CPE) usually develops within 3 days but multiple passages of five days each are recommended before concluding that a sample is NiV negative. Monolayers are examined for the presence of syncytia after incubation for 24–48 hours at 37°C. Henipavirus-induced syncytia are characterised by presence of large multinucleated cells containing viral antigen. In absence of CPE, two 5-day additional passages are recommended to confirm negative results. Immunostaining or virus neutralization tests (plaque reduction, microtitre neutralization, immune plaque assay) are applied to characterize the virus isolate and differentiate cross reactivity within Henipaviruses.
_
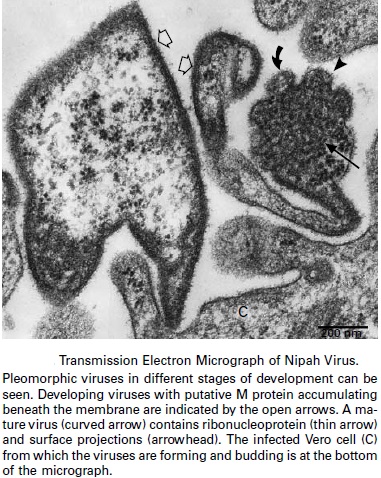
_
In early March 1999, Vero cells inoculated with CSF specimens from 3 patients with fatal cases of encephalitis developed syncytia within 5 days. The infected cells did not react with antibodies to known paramyxoviruses or other encephalitic viruses, including JE virus. However, these cells stained positively with antibodies against Hendra virus on indirect immunofluorescence assay. Cross-neutralization studies found an 8–16- fold difference in levels of antibodies to this virus, named “Nipah virus,” and levels of antibodies to Hendra viruses, which indicates that the viruses, although related, are not identical. Electron microscopic (EM) studies of the Nipah virus demonstrated features characteristic of the family Paramyxoviridae. Viruses of this family typically possess a single-stranded nonsegmental RNA genome of negative polarity that is fully encapsidated by protein. Virus particles vary in size from 120 to 500 nm. Thin-section EM studies of infected cells revealed filamentous nucleocapsids within cytoplasmic inclusions and incorporated into virions budding from the plasma membrane. Typical “herringbone” nucleocapsid structures were observed by means of negative-stain preparations. In initial reverse-transcriptase PCR experiments, the entire nucleoprotein (N) gene and 700 nucleotides from the 3 terminus of the phosphoprotein (P) gene were amplified. Nipah virus differed from Hendra virus by 21% and 25% at the nucleotide level in the N and P genes, respectively. The predicted sequence of the N protein of Nipah virus differed from that of Hendra at 42 amino acid positions (8.0%). These findings demonstrate that Hendra virus and Nipah virus represent a unique genus within the Paramyxoviridae family. This is further supported by a study that showed that the Nipah virus genome is closely related to that of Hendra virus.
_
Immunohistochemistry (IHC) can be performed to detect NiV. The N protein antigen is commonly targeted. Detection of phosphoprotein (P) antigen can also be used with IHC, although N protein antigen is expressed in greater quantities than P protein antigen and is therefore of better diagnostic value. Immunohistochemistry can be applied on formalin-fixed tissues or formalin-fixed cells of vascular endothelium from brain, lung, mediastinal lymph nodes, spleen, kidney, uterus, placenta and foetus, using antisera to NiV, rabbit antisera to plaque-purified NiV or biotinstreptavidin peroxidase-linked detection system. Immunohistochemistry can be performed on preserved tissues allowing a diagnosis to be made retrospectively. It also has the advantage that testing can be done in the absence of a BSL-4 facility. A range of polycloncal and monoclonal antisera are used, but they are not available commercially. In patients who have died, immunohistochemistry can be used to detect viral antigens in tissues. Nipah virus antigens are most likely to be found in the central nervous system (CNS), followed by the lung or kidney.
Immunofluorescence can rapidly detect NiV but cannot differentiate between henipaviruses, since mono-specific antisera to individual proteins of NiV will cross react with HeV. Negative contrast electron microscopy can be used to identify viral particles. Two monoclonal antibodies (MAb) with affinity for the N protein or P, V, and W protein of henipaviruses have been developed. MAb 1A11 C1 can detect the N protein of both NiV and HeV, while MAb 2B10 p4 can detect HeV antigen better than NiV antigen, making it possible to use 2B10 p4 for differentiation between NiV and HeV. To differentiate the two henipaviruses using immunofluorescence assays, reactivity to NiV or HeV specific antisera must be compared to positive controls of NiV or HeV. Anti-HeV antiserum neutralizes HeV at a four-fold greater dilution than it neutralizes NiV, and anti-NiV antiserum neutralizes NiV four times more efficiently than it neutralizes HeV.
_
Polymerase Chain Reaction (PCR) assay and real-time PCR can be applied with the advantage of not propagating live infectious virus. RT PCRs can be used for detection of viral sequences in CSF, throat swab or urine specimens. PCR methods and sequencing are necessary for genetic characterization of these viruses, especially with a suspected new outbreak in a geographically distinct area, as occurred in the Bangladesh outbreak (Harcourt, 2005). Nested primers coding for the M or N genes are most commonly used at present, although primers for the P gene were used in the initial outbreak in Malaysia and Singapore (Chua et al., 2000a; Daniels et al., 2001). Quantitative real-time PCR (qRT-PCR) primers and probes have been developed for the nucleocapsid (N) gene of NiV. Primers for conventional PCR and sequencing of the matrix (M) gene have also been described. Specific qRT-PCR primers and probes for the N gene of bNiV and mNiV have been described.
_
Serologic methods include neutralization tests and enzyme-linked immunosorbent assays (ELISA). Serum Neutralisation (SN) test is designated as the reference standard for anti-Henipavirus antibody detection. Cultures are read at 3 days, and those sera that completely block development of CPE are designated as positive. Immune plaque assay is an option in case of cytotoxicity. The serum neutralization test is the accepted standard for serology, but it requires BSL-4 facilities, as cell cultures must be used to determine whether a cytopathic effect has occurred. Virus neutralization tests (VNT) have been developed for high-throughput screening in BSL-2 diagnostic laboratories using recombinant vesicular stomatitis virus (rVSV) expressing NiV fusion (F) protein and receptor-binding (G). A VNT utilizing secreted alkaline phosphatase (SEAP) has been developed to detect antibodies to NiV. Using 75% reduction as the cut-off threshold, this VNT gave the same diagnostic results as a live NiV VNT performed in a BSL-4 lab. Another VNT with similar efficacy to a live NiV VNT test has been described using a luciferase reporter gene to detect neutralization of NiV proteins.
The ELISA utilizes both indirect formats for IgG and antibody capture for detecting IgM antibodies for Hendra and Nipah viruses. To perform these tests, preparation of viral antigen has been used, but research is being done to express antigen utilizing individual viral proteins such as the N protein of both viruses (Bellini et al., 2002). For the human sera, the NiV-N protein-based indirect IgG ELISA had a sensitivity of 98.6% and a specificity of 98.4%, and the NiV-N protein-based IgM capture ELISA had a sensitivity of 91.7% and a specificity of 91.8%, with reference to the CDC ELISA systems. The NiV-N-based IgM ELISA was found to be more sensitive than the inactivated-virus-based ELISA. These newer techniques allow ELISA preparation and testing to be done at facilities with BSL-2. This gives ELISA the advantage of being able to quickly detect antigen in a wider range of laboratory settings; the test is also more useful for rapid diagnosis for many cases in a suspected outbreak setting compared to serum neutralization. The recombinant G protein-based NiV ELISA is suitable for routine diagnosis of NiV and also for epidemiological surveys as it allows highly reliable testing of a large number of sera rapidly. However, the sensitivity and specificity of the ELISA test are slightly inferior to serum neutralization tests. In surveillance programs, a positive test would need to be followed with a positive virus neutralization result to confirm the diagnosis and prevent a costly and unnecessary response to a false positive result. An initial IgM anti-Nipah virus antibody response was noted in about half of patients on day one of symptoms, rising to 100% from day 3–9 (Ramasundrum et al., 2000). Over half of patients exhibited positive IgG antibody after 2 weeks, and all became positive by day 17–25. However, the presence of antibody in serum or CSF did not influence the rate of isolation of virus from CSF, nor did it correlate with decreases in morbidity or mortality (Ramasundrum et al., 1999; Chua et al., 2000). Previous NiV outbreaks have mainly relied on either IgM serology or RT-PCR testing for detection of acute infection. Studies have shown that some patients confirmed by RT-PCR or viral isolation of NiV may not have detectable IgM. Viral isolation is only performed in BSL-4 facilities.
_
Multiplexed microsphere immunoassays have been developed to detect either antibody binding to recombinant soluble NiV or HeV G protein, or antibody inhibition of ephrin-B2 receptor binding. Spectrally distinct microspheres allow specific and sensitive quantification and differentiation between HeV and NiV antibodies in a sample. Because of the recombinant subunit proteins used, this assay does not have to be conducted in a BSL4 lab if the samples are treated to inactivate virus.
_
Viral antigen capture ELISAs would also provide a high throughput format at relatively low cost for screening of the samples. Chiang et al. have reported the development of monoclonal antibody based antigen capture ELISAs for virus detection and for differentiation between NiV and HeV. Polyclonal antibody derived from the immunization of rabbits with NiV-G protein DNA vaccine construct was used for development for a novel antigen-capture sandwich ELISA system. It is suggested that considering the recent emergence of genetic variants of Henipaviruses and the resultant problems that arise for PCR-based detection, this system could serve as an alternative rapid diagnostic and detection assay.
______
Brief summary of lab diagnostic tests developed in various laboratories:
| Technique/product developed | Place |
| 1. Rapid immune plaque assay for the detection of Nipah viruse and anti-virus antibodies | CSIRO, Australia |
| 2. Solid-phase blocking ELISA for detection of antibodies to Nipah virus. | DVS, Malaysia |
| 3. Real-time RT-PCR (TaqMan) | Institute Pasteur, France |
| 4. MAb against formalin-inactivated NiV | National Institute of Animal Health Japan. |
| 5. Recombinant nucleocapsid protein produced in Escherichia coli | University Putra Malaysia |
| 6. MAb-based immunohistochemical diagnosis NiV | National Institute of Animal Health, Japan |
| 7. Recombinant glycoprotein produced in insect cells | University Putra Malaysia, |
| 8. Recombinant nucleocapsid protein produced in insect cells | University Putra Malaysia, |
| 9. Recombinant glycoprotein produced in E. coli | University Putra Malaysia |
| 10. Monoclonal antibodies against NiV (4 MAbs against “N” protein and 1 against “M” protein | NCFAD, Canada |
| 11. Indirect ELISA for the detection of Henipavirus antibodies based on a recombinant nucleocapsid protein expressed in E. coli | Chinese National Diagnostic Center for Exotic Animal Diseases, |
| 12. Indirect IgG ELISA for human and swine sera and an IgM capture-ELISA for human sera using the recombinant NiV-N protein as an antigen | Institute of Tropical Medicine, Japan |
| 13. Neutralization assays for differential Henipavirus serology using Bio-Plex Protein Array Systems | CSIRO, Australia |
| 14. Duplex nested RT-PCR for detection of Nipah virus RNA from urine specimens of bats | Chulalongkorn University Hospital, Thailand |
| 15. Monoclonal antibodies against the nucleocapsid proteins | Institute of Veterinary Sciences, China |
| 16. Neutralization test for specific detection of NiV antibodies using pseudotyped VSV | National Inst. Inf. Diseases, Japan |
| 17. Recombinant matrix protein produced in E. coli | University Putra Malaysia Malaysia |
| 18. Neutralization assay using VSV pseudotype particles expressing the F and G proteins of NiV (pVSV-NiV-F/G) as target antigens | CDC, Atlanta |
| 19. MAb based antigen capture ELISAs for virus detection and differentiation between NiV and HeV | CDC, Atlanta |
| 20. Antigen capture ELISA using polyclonal antibodies obtained by DNA immunization | National Institute Inf. Diseases, Japan |
| 21. Second generation of pseudotype-based serum neutralization assay for NiV antibodies | National Institute Inf Diseases, Japan |
_______
_______
Imaging diagnosis of nipah:
_
Chest radiograph:
Chest X-ray is abnormal in a varying number of patients, ranging from 6% to 72% in the Malaysia and Singapore outbreaks, consisting of mild interstitial infiltrates or alveolar consolidation in one or both lung fields (Paton et al., 1999; Chong et al., 2000; Goh et al., 2000). Chest radiographs were performed for 5 patients from the Faridpur outbreak; all had a history of cough and difficulty breathing, and radiographic findings for all 5 were consistent with acute respiratory distress syndrome (figure below). In patients with chest radiograph-confirmed acute respiratory distress syndrome, respiratory symptoms (cough and difficulty breathing) started within 2–5 days after the onset of fever. All 5 patients with acute respiratory distress syndrome died.
Figure above shows chest radiographs for 4 patients, showing diffuse bilateral opacities covering majority of the lung fields, consistent with acute respiratory distress syndrome.
_
CT and MRI for nipah encephalitis:
Most of the computed tomography (CT) scans of the brain did not reveal any abnormality. However, MRI abnormalities were unique, often showing multiple, small (less than 5 mm), asymmetric focal lesions in the subcortical and deep white matter without surrounding edema. These were in stark contrast to the bilateral thalamic and basal ganglia involvement in neuroimaging of JE. Even seropositive patients who were asymptomatic had white matter changes on MRI of the brain. Pathology studies indicated that underlying microinfarction might be responsible for these lesions. There was no correlation with the focal neurologic signs, depth of coma, and outcome of the patients. The lesions were attributed to widespread microinfarctions from underlying vasculitis of cerebral small vessels. MRI findings on T1- weighted imaging include multiple widespread small lesions in the white matter, mostly in the frontal and parietal lobes (Lee et al., 1999; Goh et al., 2000). The pons and cerebellum have also been affected (Lim et al., 2002). T2-weighted imaging demonstrates hyperintense lesions in gray matter and on fluid-attenuated inversion recovery sequences.
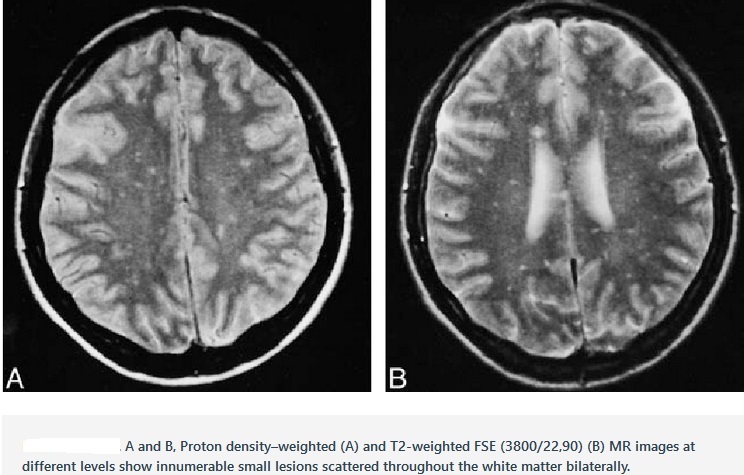
_
_
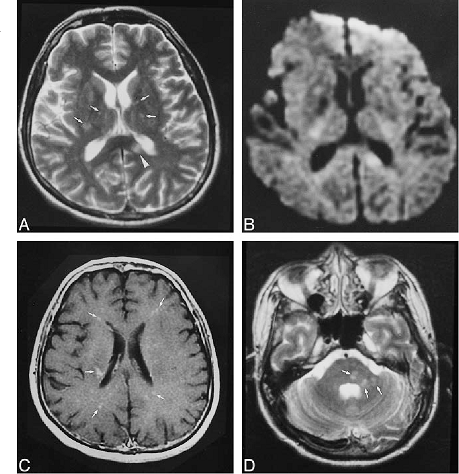
A, T2-weighted MR image shows a lesion in the corpus callosum (arrowhead) and several smaller punctate hyperintensities (arrows).
B, Only the larger lesion in the corpus callosum is visible on corresponding diffusion-weighted image.
C, Axial contrast-enhanced T1-weighted image shows several foci of enhancement (arrows).
D, Hyperintensities are seen in the pons and cerebellar peduncle (arrows) on T2-weighted image.
_
Magnetic resonance of the brain is helpful in differentiating Nipah encephalitis from other encephalitis as well as in defining between acute and late onset or a relapsed form of the disease. Features found on MR imaging in relapsed and late-onset encephalitis differed from the features in acute encephalitis in that confluent cortical involvement was the prominent finding in the former, as opposed to discrete focal lesions in the subcortical and deep white matter in the latter.
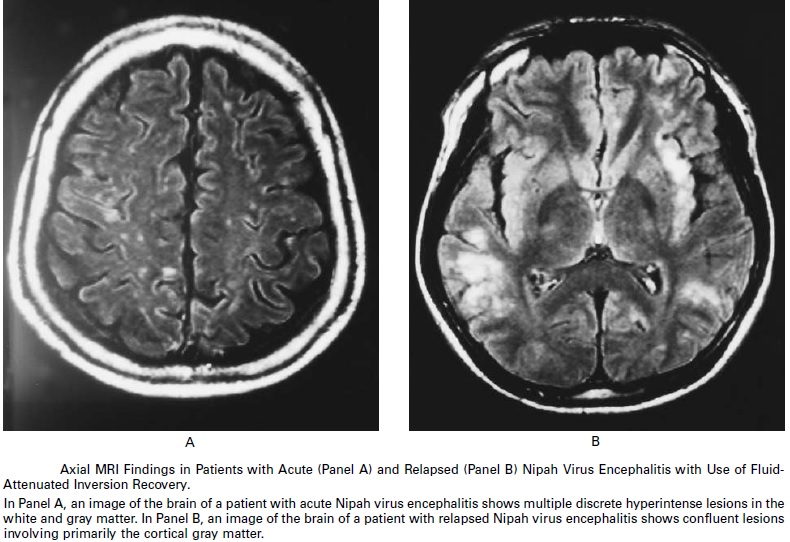
____
EEG:
Sixty-three electroencephalograms were obtained in 36 patients. The main abnormalities were diffuse slow waves with focal sharp waves (56 percent); continuous, diffuse, irregular slow waves (31 percent); and intermittent, diffuse slow waves (11 percent). Focal abnormalities were mainly seen in the temporal regions (75 percent). The electroencephalographic findings were correlated with the severity of the disease. Bilateral temporal periodic complexes of sharp and slow waves every one to two seconds were seen in deeply comatose patients; all such patients died. There was no relation between the electroencephalographic findings and the presence of myoclonus or focal lesions on neuroimaging.
_____
_____
Differential diagnosis of nipah encephalitis:
- Other viral encephalitides e.g. Herpes simplex encephalitis, Japanese Encephalitis (JE)
- Bacterial meningitis
- Cerebral Malaria
Three cases of Japanese Encephalitis have been detected from the suspected Nipah virus infection samples that were sent to the National Institute of Virology in Pune in May 2018. So Japanese Encephalitis virus should also be considered for diagnosis while screening for Nipah virus infection.
_
Differential diagnosis of Nipah virus, Japanese encephalitis and Herpes Simplex Encephalitis:
| Characteristics | Nipah virus | Japanese encephalitis | Herpes Simplex encephalitis |
| Agent | Nipah virus
(Paramyxovirus family) |
JE virus
(RNA, Flavivirus) |
HSV |
| Incubation Period | Median 10 days (range :2-21 days ) | 1-6 days, max. 14 days | 2-12 days, mean 4 days |
| Transmission | Drinking raw date palm sap, human-to-human, (close physical contact with Nipah case), animal (pig) to man | Culex mosquito (vector), human-to-human not reported | Human-human, respiratory, droplet |
| Site of involvement | Cortico-subcortical areas of cerebrum /cerebellum, brain stem | Thalamus, cortex, cerebellum, AHC | Fronto-temporal area |
| Clinical feature | Fever, headache, altered sensorium but specially associated with segmental myoclonus & respiratory involvement | Fever, headache, altered sensorium (100%) followed by convulsions and meningeal sign, abnormal movements | Same as JE but typically associated with a constellation of frontotemporal features with aphasia or mutism, personality change, and focal or generalized seizures |
| Serology / PCR | IgM /IgG (ELISA),PCR | Ag /Ab in blood /CSF | CSF PCR for
HSV DNA is diagnostic |
| CSF | Pleocytosis (10-60 cells /mm3), Protein (30- 60 mg/L), Normal glucose | Pleocytosis (10-90 cells/mm3) , Protein (900mg/L), Normal glucose | Lymphocytic pleiocytosis (typically 10–200 cells/ mm3), normal glucose, and increased protein (0.6 to 6 g/l). Red blood cells and xanthochromia may be present |
_____
_____
Sample collection and laboratory biosafety vis-à-vis nipah:
Nipah virus being a BSL-4 agent, universal, standard droplet and bio-containment precautions should be followed during contact with excretions, secretions and body fluids of suspected patient. Adequate biosafety precautions should be adopted during collection/transport/ storage/ processing of clinical samples from suspected cases. The samples should be collected as early as possible (preferably within 4-5 days on onset of illness) with all biosafety precautions and accompanied with detailed history of patients on the performa which can be obtained from the testing laboratory. During sample collection wear complete disposable Personal Protective Equipments (N 95 mask, double surgical gloves, gowns, goggles etc.). Wash hands with soap and water at least for 30 seconds and then clean hand using 1-2 ml of alcohol based hand sanitizer before and after collection of samples.
The samples may be as follows
- Throat swab to be collected in viral transport medium
- Urine approx 10 ml in universal sterile container
- Blood in plain vial (at least 5ml)
- CSF (at least 1 ml) in a sterile container
Samples should be safely packed in triple container packing and should be transported under cold chain (2-8°C) to the testing laboratory with prior intimation. Before dispatching the sample, disinfect the outer surface of container using 1:100 dilution of bleach or 5% Lysol solution. Sample containing vials should be kept in good quality plastic bags tied with rubber bands so that inside material if leaks should not come out of bag. The plastic bag should be kept in another container which should be sealed with adhesive tape. This carrier should be placed in another plastic bag sealed with rubber bands and placed in thermocol/vaccine carrier containing ice. The case sheets with complete information should be placed in plastic bag and should be pasted outside the container. Samples should be transported at 2-8°C if they arrive at the laboratory with 48 hours; if shipping time is expected more than 48 hours, the samples should be sent using dry ice. Samples should not be held at -20°C for long periods. The sample must be stored at – 70°C if storage is required for longer periods.
__
Laboratory biosafety:
“Laboratory bio-safety” is the term used to describe the containment principles, technologies and practices that are implemented to prevent unintentional exposure to pathogens and toxins, or their accidental release.
“Laboratory biosecurity” refers to institutional and personal security measures designed to prevent the loss, theft, misuse, diversion or intentional release of pathogens and toxins. Bio-safety protection is to protect laboratory workers, clinical specimens and the environment. Diagnostic and health-care laboratories (public health, clinical or hospital-based) must all be designed for at least Bio-safety Level 2 or above if required. As no laboratory has complete control over the specimens it receives, standard precautions should always be adopted and practiced. The basic objective of a biosafety program is the containment of potentially harmful biological agents. The purpose of containment is to reduce or eliminate exposure of laboratory workers, other persons, and the outside environment to potentially hazardous agents. The use of vaccines may provide an increased level of personal protection. The term “containment” is used in describing safe methods, facilities and equipment for managing infectious materials in the laboratory environment where they are being handled or maintained. The appropriate combination of the elements of containment required in a laboratory is determined on the basis of the risk assessment of the work to be done with a specific agent.
_
Biosafety Levels
Four Biosafety Levels (BSLs) are described based on combinations of laboratory practices and techniques, safety equipment, and laboratory facilities. Each combination is specifically appropriate for the type of procedures performed and documented or suspected routes of transmission of infectious agents. BSL-1 are basic teaching laboratories, BSL-2 are diagnostic services and research laboratories, BSL-3 are high containment special diagnostic services laboratories and BSL-4 are maximum containment laboratories to handle dangerous pathogens. Laboratory biosafety manual (3rd Ed), WHO, describes relative hazards of infective microorganisms by risk groups for laboratory work. The classification of microbiological agents is based on their association with, and resulting severity of, disease in humans. The risk group of an agent should be one factor considered in association with mode of transmission, procedural protocols, experience of staff, and other factors in determining the BSL in which the work will be conducted.
Classification of infective microorganisms by risk group (WHO Laboratory Biosafety Manual, 3rd Ed, 2004)
- Risk Group 1 (no or low individual and community risk)
A microorganism that is unlikely to cause human or animal disease.
- Risk Group 2 (moderate individual risk, low community risk)
A pathogen that can cause human or animal disease but is unlikely to be a serious hazard to laboratory workers, the community, livestock or the environment. Laboratory exposures may cause serious infection, but effective treatment and preventive measures are available and the risk of spread of infection is limited.
- Risk Group 3 (high individual risk, low community risk)
A pathogen that usually causes serious human or animal disease but does not ordinarily spread from one infected individual to another. Effective treatment and preventive measures are available.
- Risk Group 4 (high individual and community risk)
A pathogen that usually causes serious human or animal disease and that can be readily transmitted from one individual to another, directly or indirectly. Effective treatment and preventive measures are not usually available.
Note:
Health care workers and pathologists have been documented to have been accidentally exposed to Nipah virus, but after stringent testing, none were shown to have contracted the virus. This proves that close contact with nipah patient leads to human transmission and not accidental needle prick.
_____
_____
Treatment of nipah virus disease:
Currently there is no effective treatment for Nipah virus infection. The treatment is limited to supportive care. It is important to practice standard infection control practices and proper barrier nursing techniques to avoid the transmission of the infection from person to person. All suspected cases of Nipah virus infection should be isolated and given intensive supportive care. Ribavirin has been shown to be effective in Malaysia outbreak. Passive immunization using a human monoclonal antibody that targets the Nipah G glycoprotein has been evaluated in the ferret model as post-exposure prophylaxis. m102.4, a human monoclonal antibody, has been used in people on a compassionate use basis in Australia and is presently in pre-clinical development. The anti-malarial drug chloroquine was shown to block the critical functions needed for maturation of Nipah virus, although no clinical benefit has yet been observed.
_____
Antiviral agents:
Ribavirin is a well-known first line treatment strategy for suspected viral infections of unknown etiology. Ribavirin exhibits antiviral activity against a wide variety of both RNA and some DNA viruses (Sidwell et al., 1972) and is an accepted or approved treatment for several viral infections including respiratory syncytial virus and arenaviral hemorrhagic-fevers (reviewed in (Snell, 2001)). In vitro studies have shown that ribavirin is effective against both Hendra and Nipah virus replication (Aljofan et al., 2009; Wright et al., 2005). Ribavirin and acyclovir have been used to treat Nipah virus infection. In Malaysia, ribavirin was administered orally or intravenously to 140 persons with Nipah virus encephalitis and compared to a group of 54 control patients who did not receive ribavirin. A total of 45 deaths in the treated group (32%) compared to 29 deaths in the control group (54%) suggested a 36% reduction in mortality with ribavirin administration (Chong et al., 2001), (P = 0.011). Duration of ventilation and total hospital stay were both significantly shorter in the ribavirin group (P = 0.0002 and <0.0001, respectively). There was a nonsignificant trend to a reduction in neurological deficits in survivors who received ribavirin (P = 0.17). This study used decremental doses of oral or intravenous ribavirin (beginning with 2 g orally or 30 mg/kg iv) over a week to 10 days. There is certainly a paucity of blinded, randomized, controlled trial data on the efficacy of ribavirin in this condition, but it is clearly quite incorrect to say that ribavirin was ineffective. In Singapore, acyclovir was administered to all encephalitis patients during the Nipah outbreak (Paton et al., 1999; Bellini et al., 2002). Only one fatality occurred in Singapore, but the effect that the drug had on the course of disease is unclear. However, more recent animal studies have revealed no therapeutic benefit of either drug. Two studies in hamsters and one study in nonhuman primates (African green monkey (AGM)) showed that ribavirin treatment only delayed but did not prevent death after Nipah or Hendra virus infection (Freiberg et al., 2010; Georges-Courbot et al., 2006; Rockx et al., 2010) and AGMs treated with ribavirin following Hendra virus infection had marked increases of neurological symptoms.
_
Favipiravir (T-705) is a purine analogue antiviral approved for use in Japan against emerging influenza strains; and several phase 2 and 3 clinical trials are ongoing in the United States and Europe. Favipiravir has demonstrated efficacy against a broad spectrum of RNA viruses, including members of the Paramyxoviridae, Filoviridae, Arenaviridae families, and the Bunyavirales order. A study demonstrates that favipiravir has potent antiviral activity against henipaviruses. In vitro, favipiravir inhibited Nipah and Hendra virus replication and transcription at micromolar concentrations. In the Syrian hamster model, either twice daily oral or once daily subcutaneous administration of favipiravir for 14 days fully protected animals challenged with a lethal dose of Nipah virus. This first successful treatment of henipavirus infection in vivo with a small molecule drug suggests that favipiravir should be further evaluated as an antiviral treatment option for henipavirus infections.
_
Recently, the adenosine nucleoside analogue GS-441524, and its monophosphate prodrug GS-5734, were demonstrated to have in vitro antiviral activity against NiV and HeV with EC50 values between 0.49 to 1 M and 0.032 to 0.055 M, respectively. Importantly, GS-5734 was protective in a non-human primate model for Ebola virus post exposure and is currently in phase 2 clinical development for the treatment of Ebola virus disease. Additionally, another nucleoside analogue, R1479 (balapiravir), demonstrated in vitro antiviral efficacy against NiV and HeV with EC50 values of 4M and 2.25M, respectively.
_
Hendra and Nipah viruses were shown to be susceptible to chloroquine in 2009 study:
The researchers, based in Weill Cornell’s pediatrics department, were surprised by their discovery that chloroquine, a safe, low-cost agent that has been used to combat malaria for more than 50 years, is a highly active inhibitor of infection by Hendra and Nipah. Chloroquine does not prevent Hendra or Nipah virus from entering the cell. Instead, the chloroquine molecule appears to block the action of a key enzyme, called cathepsin L, which is essential to the virus’s growth and maturation. Without this enzyme, newly formed Hendra or Nipah viruses cannot process the protein that permits the viruses to fuse with the host cell. Newly formed viruses then cannot spread the infection; in other words, they can invade, but cannot cause disease. Several other zoonotic viruses also depend on cathepsin L most notably, Ebola. However, chloroquine was unable to prevent Nipah infection or disease in ferrets (Pallister et al., 2009). Several studies using disease-relevant animal models have repeatedly demonstrated ribavirin monotherapy as well as combination treatment with chloroquine to be ineffective at reducing the mortality of henipavirus infections.
______
Passive immunotherapy for nipah:
Hendra and Nipah virus attach to host cell-surface displayed ephrin-B2 or -B3 proteins and infect host cells by the coordinated activity of their attachment (G) and fusion (F) glycoproteins. The G glycoprotein monomer consists of a stalk and globular head (figure below) and the atomic structures of both the Nipah and Hendra virus G glycoprotein’s globular head domain have been determined alone and in complex with ephrin proteins. The F glycoprotein mediates the membrane fusion process between the viral and host cell membranes by a Class I fusion mechanism that is initiated following the G glycoprotein engagement of ephrin receptor. The susceptible host species and associated cellular tropism and pathology of Hendra and Nipah virus has in large part been explained by their use of the highly conserved ephrin-B2 and -B3 proteins as entry receptors. In addition and of importance to countermeasure development, the henipavirus G and F envelope glycoprotein spikes are major targets of virus-neutralizing antibodies and as discussed below, the development of potential vaccines have largely focused on these important structural components of the virion. A neutralizing human monoclonal antibody (hmAb), m102.4 recognizes the receptor binding domain of the HeV and NiV G glycoproteins. This hmAb potently neutralized both viruses in vitro and maintained its biological activity in vivo suggesting its possible utility as a passive therapeutic modality following henipavirus infection .
_
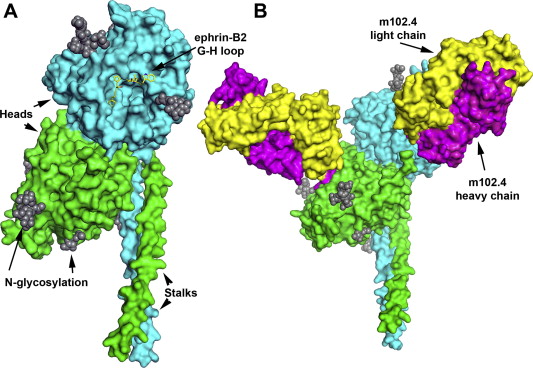
Figure above shows Model of the Hendra virus soluble G glycoprotein subunit vaccine (HeV-sG) and its complex with the henipavirus-neutralizing human monoclonal antibody m1024. (A) The HeV-sG glycoprotein subunit vaccine is composed of the entire ectodomain (amino acids 76–604) of the HeV G glycoprotein. Here, HeV-sG is shown as dimer with one monomer colored green and the other cyan. The secondary structure elements of the two globular head domains are derived from the crystal structure of the HeV G head domain (Colgrave et al., 2011; Xu et al., 2012b), and the stalk regions of each G monomer (residues 77–136) are modeled (Kelley and Sternberg, 2009). N-linked glycosylation sites shown as gray spheres. The ephrin-binding face of the cyan globular head is facing forward with an overlay of the interacting ephrin-B2 G-H loop residues in yellow. (B) The HeV-sG dimer shown in complex with two m102.4 Fab antibody fragments. The two HeV-sG monomers are colored green and cyan as in panel A and rotated slightly to the right. The two Fab m102.4 molecules are shown with their heavy chains colored in magenta and light chain – in yellow, each binding one globular head domains of G. (Xu, K. et al., submitted).
__
Passive immunotherapy with polyclonal or monoclonal antibody specific for the viral envelope glycoproteins has proved successful from initial proof-of-concept findings from several studies carried out in hamsters (Guillaume et al., 2004, 2006). Presently, the only reported and effective post-exposure therapy against Hendra or Nipah virus infection and one that could likely be approved in the near future for use in people has been a human monoclonal antibody (mAb) known as m102.4 which was isolated from a recombinant naïve human phage-displayed Fab library (Zhu et al., 2008). The m102.4 mAb has exceptionally potent neutralizing activity against both Nipah and Hendra viruses and its epitope maps to the ephrin receptor binding site (figure above). Testing of m102.4 has confirmed its neutralization activity against several isolates; NiV-Malaysia, HeV-1994, HeV-Redlands, NiV-Bangladesh (Bossart et al., 2009). Effective post-exposure efficacy with m102.4 has now been demonstrated in both ferrets and nonhuman primates (African green monkey (AGM)) infected with either Hendra virus or Nipah virus (see table below). The successful m102.4 passive immunotherapy in the AGM was recently reported in a study designed to reflect a possible real life scenario requiring mAb as a post-exposure treatment, and was a follow-up from the initial successful m102.4 post-exposure therapy carried out in ferrets (Bossart et al., 2009). Fourteen monkeys were challenged intratracheally with Hendra virus and 12 animals were infused twice with a 100 mg dose (∼20 mg/kg) of m102.4 beginning at 10 h, 24 h or 72 h p.i. with the second infusion ∼48 h later. All 12 animals that received m102.4 survived infection; whereas the untreated control subjects succumbed to severe systemic disease by day 8 (Bossart et al., 2011). There was no evidence of Hendra virus mediated pathology in any of the m102.4-treated animals and no infectious Hendra virus could be recovered from any tissues from any m102.4-treated subjects.
_
Evaluation of post-exposure therapy with the henipavirus G glycoprotein-specific human monoclonal antibody m102.4 against lethal henipavirus challenge in laboratory animals:
| Virus | Animal model | Experimental design and results |
| Hendra | AGM | A 10, 24, or 72 h post-exposure use of m102.4 could protect against a 10-fold lethal intratracheal virus challenge |
| Ferret | A 10 or 24 h post-exposure dose of m102.4 could protect against a 10-fold lethal oronasal virus challenge | |
| Nipah | Ferret | A 10 h post-exposure dose of m102.4 could protect against a 10-fold lethal oronasal virus challenge |
| AGM | A 24, 72, or 120 h post-exposure use of m102.4 could protect against a 10-fold lethal intratracheal virus challenge |
The administration of m102.4 mAb was performed by infusion in all studies with the exception of one ferret in one study were it had to be administered by intraperitoneal injection (Bossart et al., 2009). All studies to date have reported that all animals receiving m102.4 post-exposure therapy have survived virus challenge, some with varying levels of mild to moderate illness. The hmAb m102.4 is an ideal therapeutic, targeting a critical functional domain of the G glycoprotein, and is a fully human IgG.
______
Development of therapeutics & treatment modalities:
Monoclonal antibodies tend to be quite expensive and are very disease specific. In addition to furthering the antibody therapeutic studies, future progress should focus on siRNAs, small-molecule drugs, the screening of currently approved drugs for NiV efficacy and, most especially, identifying broad-spectrum antivirals. The basic science work has already begun to identify novel targets for future drug development including viral protein motifs, viral RNAs, host proteins involved in antiviral responses and proteins hijacked by NiV, as well as host RNA targets. The ability to target host proteins or RNAs has the advantage of making resistance development much more difficult and is much more likely to offer potential for broad-spectrum antiviral activity as these pathways are often utilized by many different viruses.
The last 5 years have shown light on the importance of the NiV Matrix protein, Phosphoprotein, and the accessory proteins V, W and C in the pathogenesis of NiV in small animal models. Studies in the near future will confirm if these interactions are also important in pathogenesis in nonhuman primates. These will likely be the important viral targets, along with the outer membrane Glycoprotein and Fusion protein, for novel drug development. In addition, studies in the near future will also further characterize the interactions between these NiV proteins and specific host proteins to identify the exact viral motifs and/or host protein motifs to target in future drug development.
An additional area of importance that will shine light on human immune factors that can protect against NiV infection will involve analysis of the Cynomolgus macaques, African green monkey and squirrel monkey genomes. Cynomolgus macaques do not develop signs of NiV disease while African green monkeys and squirrel monkeys are susceptible to NiV infection. Cynomolgus macaques are found in Southeast Asia where NiV is endemic and likely co-evolved with NiV or related henipaviruses. African green monkeys, on the other hand, are from Africa and squirrel monkeys are from South and Central America. NiV is not endemic in these places and thus these species are naive to exposure to NiV. As DNA sequencing technology continues to improve and become cheaper, soon the genomes from these primate species will be fully sequenced and annotated and this will allow for experiments to understand the host genetics that allow some primate species to be susceptible while others are not. This understanding will offer additional human targets for therapeutic intervention.
Experiments are also underway examining nonpharmacological treatment options for treating animals infected with NiV. With high-level supportive care, such as found in a hospital intensive care unit, most patients are able to survive the respiratory component of NiV, but the meningoencephalitic component often leads to death with many survivors having long-term neurological sequelae. The biosafety level 4 laboratory at the United States National Institute of Allergy and Infectious Diseases Integrated Research Facility now has the capacity to handle nonhuman primates in a setting with advanced medical imaging and intensive medical care, thus allowing for experiments examining means of decreasing morbidity and mortality in NiV infection using methods other than novel drugs. Alternatively, there is a new ferret model with a recombinant NiV mutant lacking both C and W protein expression that drastically decreases the respiratory component of NiV disease while still allowing for neurological disease which can lead to death or survival with long-term neurological sequelae, similar to human cases. This model will also allow for experiments in means of decreasing morbidity and mortality in NiV infection using similar alternative methods in laboratories not equipped with the advanced medical facilities of the Integrated Research Facility.
______
______
Relapse, complications and mortality of nipah virus disease:
Most people who survive acute encephalitis make a full recovery, but long term neurologic conditions have been reported in survivors. Approximately 20% of patients are left with residual neurological consequences such as seizure disorder and personality changes. A small number of people who recover subsequently relapse or develop delayed onset encephalitis. In survivors, there is a potential for relapse months to years after infection, relapse occurs in approx. 3 to 8%. The case fatality rate in humans is estimated at 40% to 75%. This rate can vary by outbreak depending on local capabilities for epidemiological surveillance and clinical management.
_
Relapse occurs in 3–8% of patients, occasionally causing more severe clinical symptoms than the initial manifestation (Goh et al., 2000; Tan et al., 2002). Relapse occurs at a mean of 8 months after initial presentation. Late-onset encephalitis occurs in 3% of infected patients who were initially asymptomatic or non-encephalitic, and has been reported to occur as late as 4 months after initial infection (Goh et al., 2000; Wong et al., 2001). Clinical symptoms of relapsed and late-onset encephalitis are similar to those of initial encephalitis. Patients who had recovered from clinical illness and some who had only serological evidence of infection but were not ill were monitored for up to 2 years. Goh et al. (2000) reported four patients who had late neurologic dysfunction, with three of them having a relapse 13–39 days after an initial mild illness. Another patient was seropositive for antibodies against Hendra virus but remained asymptomatic until he became ill 11 weeks later. He subsequently died. One had an isolated nuclear third-nerve palsy, from which she gradually recovered. Three others had altered consciousness, and two of these patients had seizures. Dysphasia and segmental myoclonus with ataxia occurred in one patient each. In three patients, the cerebrospinal fluid examination results were abnormal, with lymphocytic pleocytosis and elevated protein levels; the virus could not be isolated from the cerebrospinal fluid. In patients with relapses or disease of delayed onset, diffuse confluent involvement of the cortical gray matter was found on MRI.
__
The exact prevalence of individuals with neurologic or psychiatric sequelae of Nipah virus encephalitis is uncertain, as study results vary and sample sizes are generally small. The largest study found 15% (14 of 110 individuals) with residual neurologic deficits (Goh et al., 2000). Higher percentages of residual neurologic, psychiatric, or cognitive symptoms have been reported in smaller studies (Limet al., 2003; Ng et al., 2004). Neurologic sequelae have included residual cognitive deficits, verbal impairment, cranial nerve palsy, cerebellar abnormalities, and persistent vegetative state. Neuropsychiatric sequelae have included personality changes and major depression. Chronic fatigue syndrome has sometimes also been reported as a sequela of Nipah virus encephalitis.
_
The morbidity and mortality rates of henipaviruses have varied over the course of various outbreaks. NiV is highly contagious with high morbidity in swine, and can be as high as 100 percent depending on density of the affected population and age of the animal. Mortality is higher in piglets (40 percent) than in young swine or older animals (<5 percent). For HeV, morbidity in horses is relatively high, though the infection in horses does not appear to be highly contagious. The case fatality rate is high in both humans and horses, though an estimate of the mortality rate is hard to provide given the relatively few outbreaks that have occurred. The overall mortality rate of symptomatic Nipah virus infection in humans differs by country: 40% (105/265) in Malaysia, 9% (1/11) in Singapore, 74% (exact figures uncertain) in India, and 76% (66/87) in Bangladesh. It is uncertain whether these differences are due to virulence factors in the virus or whether they reflect the level or availability of supportive care in each country. Factors that predicted mortality included the presence of doll’s-eye reflexes, tachycardia, high fever, hypertension, and a positive viral culture of the CSF (Chong et al., 2000; Chua et al., 2000b; Goh et al., 2000). In clinical cases, the fatality rate has ranged from 38% to approximately 70-75% in various outbreaks, with higher rates reported from some small case series. The case fatality rate is reported to be much higher in Bangladesh and India than Malaysia, but whether this is due to strain variability or to differences in healthcare is uncertain. One study reported a higher mortality rate in people with diabetes.
____
Hendra and Nipah viruses are deadly:
Henipavirus, including Hendra and Nipah viruses, is a group of emerging bat-borne paramyxoviruses which were responsible for severe disease outbreaks in humans, horses and pigs. The mortality rate of human infection varies between 50 and 100%, making them one of the most deadly viruses known to infect humans. Its use of highly conserved cell surface molecules (ephrin) as entry receptors and its highly effective replication and fusion strategies are believed to be important characteristics responsible for its high pathogenicity. Henipavirus also encodes multiple accessory proteins which play a key role in evasion of host innate immune responses. Their unique genetic constitution, high virulence and wide host range set them apart from other paramyxoviruses.
______
______
Risks factors, control and prevention of nipah virus disease:
_
Risk factors:
The risk of exposure is high for hospital workers and caretakers of those infected with the virus. In Malaysia and Singapore, Nipah virus infection occurred in those with close contact to infected pigs. In Bangladesh and India, the disease has been linked to consumption of raw date palm sap (toddy) and contact with bats. Eating pork did not transmit the infection and all of the reported cases involved pig farmers, handlers, or abattoir workers. In a case control study of risk factors for the acquisition of disease, infected persons were more likely than community-farm controls to report increased numbers of sick or dying pigs on the farm. The former were also more likely than case-farm controls to perform activities requiring direct contact with pigs. Similar results were obtained in a study involving abattoir workers in Singapore. Army personnel who were involved in culling pigs in outbreak-affected states did not appear to be at significant risk. In a cross-sectional survey of 1,412 military personnel involved in culling activities, only six had detectable antibody to the Nipah virus, all of whom had reported direct contact with live pigs. Infected pigs appeared to have respiratory symptoms and their autopsies demonstrated prominent respiratory tract involvement. Pigs were later found to shed the virus in nasopharyngeal secretions, thereby infecting humans. The abnormal chest radiographs in some humans lend weight to the respiratory route of transmission. A recent study isolated the virus from the upper respiratory secretions and urine in 8 of 20 infected patients. To reduce risks of nosocomial transmission, authors advised that standard and droplet infection control practices be maintained in view of the high case-fatality rate of the disease. The isolation of the virus in respiratory secretions and urine was not associated with poor prognosis. Conversely, there was a significant co-relation between CSF isolation and mortality and clinical features that suggested a poor prognosis. The authors postulated greater viral replication in the central nervous system, resulting in the high mortality.
_____
Disinfection:
Like other paramyxoviruses, Nipah virus is readily inactivated by soaps, detergents and many disinfectants. Routine cleaning and disinfection with sodium hypochlorite or commercially available disinfectants is expected to be effective. NiV can be inactivated by 0.1% formalin and 0.5% household bleach. In general, paramyxoviruses are susceptible to acids, alcohols, aldehydes, alkalis, halogens, and oxidizing agents. Sodium hypochlorite was recommended for the disinfection of pig farms in Malaysia. The effect of heat may depend on the substrate.
_
Resistance of Henipaviruses to Physical and Chemical Action:
| Action | Resistance |
| Temperature | Other animal Paramyxoviruses are inactivated by 60ºC/60 minutes. Nipah virus concentrations decreased but the virus was not completely eliminated in artificial palm sap held at 70°C for 1 hour. However, it was completely inactivated by heating at 100°C for more than 15 minutes. Henipaviruses are rapidly inactivated by desiccation at both 22°C and 37°C. |
| pH | Stable between pH 4.0 and 10.0. |
| Chemicals/Disinfectants | Paramyxoviruses are susceptible to common soaps and disinfectants; lipid solvents (alcohol and ether) and sodium hypochlorite solutions were used effectively in outbreaks for cleaning and disinfection. |
| Survival | Survives for long periods in favorable conditions; survives for days in fruit bat urine and contaminated fruit juice. |
______
Effective control measures:
In Malaysia, the problem was tackled 19 years ago and the disease never recurred in the country, thanks to the effective way Nipah had been tackled. Over 1 million pigs were culled when Nipah virus infection was confirmed. That went a long way in controlling the spread of the virus. After Nipah virus became a concern, Malaysia introduced stringent measures to control rearing of pigs. Animal welfare department officials regularly inspected pig farms and the practice is still continued. They check the health of the animals, too. A ban was imposed on rearing pigs in open areas. This was considering the fact that pigs had got the virus from fruits partially eaten by bats while wandering in these open areas. Nipah does not spread through air or by mosquitoes. Even now, samples of blood and cerebrospinal fluid of patients suffering from encephalitis are checked for Nipah virus. Strict guidelines have been issued by the health department in this regard. In animal farms, cleanliness and hygiene should be made mandatory. However, in Bangladesh, Nipah outbreaks have recurred several times. It has been observed that bats spread the disease in Bangladesh, too. Most of the victims were children who ate fruits bitten by bats. Several health care workers who treated the children were also affected. In Bengal too, bats had spread Nipah virus in Siliguri and Nadia districts. Killing bats is not the solution because bats are an integral part of our ecosystem. Migratory bats choose certain means for perpetuation of species and other purposes. Humans should desist from venturing into their habitats.
_
Pigs seem to be important amplifying hosts for Nipah virus, and preventing infections in this species can decrease the risk of infection for humans. Sick animals should not be used for food, even if the meat is to be cooked, as the slaughter process can increase human exposure to viruses in the tissues. Close contact with fruit bats and their secretions and excretions should also be avoided. Bats have been observed visiting date palm sap collection sites at night, and can contaminate collection pots with urine and saliva. While the general recommendation is to avoid drinking any unpasteurized juices in endemic regions, keeping bats away from sap collection sites with protective coverings (e.g., bamboo sap skirts) may be helpful in areas where people are unlikely to stop drinking raw date palm sap. Smearing lime on the collection area to discourage bats appeared to have little inhibitory effect in one study. Fruit should be washed thoroughly, peeled or cooked before eating. Good personal hygiene, including hand washing, is likely to reduce the risk of infection from the environment. Nipah virus has been classified as a Hazard Group 4/ BSL4 pathogen; infected animals, body fluids and tissue samples must be handled with appropriate biosecurity precautions. People who come in close contact with potentially infected animals should wear protective clothing, impermeable gloves, masks, goggles and boots. Because Nipah virus can be transmitted from person to person, barrier nursing should be used when caring for infected patients. Patients should be isolated, and personal protective equipment such as protective clothing, gloves and masks should be used. Good hygiene and sanitation are important; in one study, hand washing helped prevent disease transmission. Vaccines are currently not available for humans.
_
Nipah and Hendra virus infections can theoretically be prevented by avoiding direct or indirect contact with fruit bats or fruit bat urine or droppings. Using precautionary measures such as gloves and masks may be considered in locations with fruit bats. Fruit or other products from trees where fruit bats roost should be carefully washed.
Because it is assumed that these viruses can spread through respiratory droplets or by contact, caution should be used when caring for an infected individual, including frequent hand washing, avoidance of direct contact with urine or salivary secretions, and wearing a mask. To prevent nosocomial transmission of the viruses, contact and droplet precautions seem prudent when taking care of any patient with suspected Nipah or Hendra virus infection.
_____
Contain the outbreak:
A single confirmed case of NiV infection constitutes an outbreak. Case investigation determines the likely source of infection through a detailed history of the patient’s work and movements. Further cases may be identified through active case finding among close contacts with similar exposure. Contact tracing, aggressive monitoring and quarantining of suspect cases are effective forms of infection control and containment. These have been appropriately implemented by the responsible government agencies responding to recent outbreak in Kerala. Educational efforts combined with preventive measures appear to be effective. Examples include:
- The use of universal precautions and appropriate PPE (gloves, masks and/or face shield) is sufficient to limit the spread of Nipah virus to patient caretakers including family relatives and healthcare workers.
- Funeral practices that avoid direct contact with the deceased can cut the chain of transmission.
- Avoiding direct contact with bodily fluids, especially respiratory secretions of infected individuals.
- Counsel relatives to avoid prolonged close contact with the infected individual (e.g. sleeping beside patient, sharing of foods, etc.). Maintain >1 full-stretched arm distance (1 metre or 3 feet) from patient.
- All the above require culturally sensitive educational campaigns targeted to the affected community.
- There is evidence that Nipah virus RNA+ patients are more likely to contaminate towels, bedsheets, and bed rails. A study also showed that Nipah virus RNA could also be detected on the surrounding walls and bedframe of a deceased Nipah virus infected patient. Thus, infection controls should also target hospital surfaces, which will reduce the risk of fomite transmission.
_____
Control of Nipah Virus in Domestic Animals:
Surveillance is an important tool for early detection for illnesses caused by the henipaviruses. Surveillance for Hendra virus illness has been established in Australia, and surveillance for encephalitic disease has been implemented in Malaysia, Thailand, and Bangladesh. Clusters of respiratory or encephalitic illness in humans or in certain animals, occurring in geographic locations where Pteropus bats are known to be endemic, should raise awareness of the possibility of henipavirus infection. Thus, surveillance in animals such as horses and pigs is also important for the early detection of Nipah and Hendra virus infections. No specific vaccine is available against Nipah or Hendra virus. However, active immunization against Nipah virus and passive transfer of antibody to Nipah virus has shown promising results in hamster models (Guillaume et al., 2004). Taking into account the human health implications, all field investigations should take necessary precautions to prevent infection. This includes prompt and accurate veterinary investigations on suspected clinical cases especially in pigs. Any respiratory or neurological conditions of swine in an area known to have pteropid bats, should consider Nipah as a rule out. Nipah should be suspected if pigs also have an unusual barking cough or if human cases of encephalitis are present. Symptoms in pigs are not dramatically different from other respiratory and neurological illnesses of pigs. Differential diagnosis should be applied in case of deaths of suckling pigs and piglets, sudden death in boars and sows, abortions and other reproductive dysfunction, respiratory diseases with harsh, non-productive coughing, and in cases with encephalitic manifestations of trembling, muscular incoordination and myoclonus leading to lateral recumbency. In pig farms contact with fruit bats and their secretions should be avoided using screens at open-air access. Control of any access to swine by other wild or domestic animals should be also ensured. Routine cleaning and disinfection of animal farms (with sodium hypochorite or other detergents) is expected to be effective in preventing infection. If an outbreak is suspected, the animal premises should be quarantined immediately. Culling of infected animals, with close supervision of burial or incineration of carcasses, may be necessary to reduce the risk of transmission to people. All materials and equipment from affected farms should be cleaned and disinfected. Restricting or banning the movement of animals from infected farms to other areas has to be applied to reduce the spread of the disease.
____
Prevention through modifying risk factors:
Preventing humans from being infected through non-vaccination efforts will perhaps play an even more important, and less expensive, role in controlling NiV disease. These efforts fall into two broad categories depending on the local epidemiology. In regions where livestock act as an intermediate host, preventing animals from coming into contact with NiV is relatively straightforward and involves keeping bat roosting trees and fruit trees away from areas of susceptible livestock farms and pastures. This effort has already been largely effective in some areas including Malaysia.
In regions where the primary source is contaminated date palm sap, this is a trickier endeavor as it involves alterations in human behavior. In Bangladesh, date palm sap is collected from taps overnight, and infrared cameras have found nocturnal evidence of these bats drinking from, and urinating and defecating in, the open jars collecting date palm sap. Efforts to modify the human behaviors and prevent NiV infection in these regions are already underway and, if given sufficient resources and support, they are likely to lead to a drastic decrease in incidence of NiV outbreaks. These efforts include campaigns to increase awareness of NiV disease and its source through written, radio and televised means; using inexpensive and often locally produced tree skirts to prevent bats from accessing the collection jars; and encouraging the boiling or pasteurization of date palm sap before consumption as this inactivates NiV. Overall, these efforts have so far met with success where implemented; however, implementation has proved difficult in many regions due to traditional cultural practices.
With climate change and human encroachment into flying fox habitats, it is likely that outbreaks will begin to occur in regions that have not yet experienced NiV outbreaks. Therefore, prevention methods will have to be adapted to the new local cultures and the importance of having a vaccine available will be even further underscored for containment in these new regions.
______
Advisory for health care personnel:
- Wash hands thoroughly with soap and water for 20 seconds after contact with a sick patient.
- While handling Nipah cases (suspected/ confirmed), standard precautions for infection control should be practiced. • For aerosol generating procedures, PPE such as individual gowns (impermeable), gloves, masks and goggles or face shields and shoe cover and the procedure should be performed in airborne isolation room.
- Dedicated medical equipment should be used (preferably disposable whenever possible).
- All non-dedicated, non-disposable medical equipment used for patient care should be cleaned and disinfected as per manufacturer’s instructions and hospital policies.
- Use of injections and sharps should be limited.
- If the use of sharp objects cannot be avoided, ensure that the following precautions are observed:
-Never replace the cap on a used needle.
-Never direct the point of a used needle towards any part of the body.
-Do not remove used needles from disposable syringes by hand, and do not bend, break or otherwise manipulate used needles by hand.
-Never re-use syringes or needles.
-Dispose of syringes, needles, scalpel blades and other sharp objects in appropriate, puncture-resistant containers.
-Ensure that containers for sharps objects are placed as close as possible to the immediate area where the objects are being used (‘point of use’) to limit the distance between use and disposal, and ensure the containers remain upright at all times.
-Ensure that the containers are securely sealed with a lid and replaced when ¾ full.
– Ensure the containers are placed in an area that is not easily accessible by visitors, particularly children (e.g. containers should not be placed on floors, or on the lower shelves of trolleys in areas where children might gain access).
-Closed, resistant shoes (e.g. boots) should be used by all individuals in the patient care area to avoid accidents with misplaced, contaminated sharp objects.
- Safe waste disposal for potentially infected material including used PPE, linen, clothing of patient according standard biomedical waste management guidelines.
- Admit all suspected cases of Nipah to the isolation ward/ facility in the hospital. Once the case is suspected of Nipah, attendants should not be permitted in the ward.
- Segregate all suspected cases of Nipah patients from all patients in the isolation ward/ facility. Maintain at least 1 meter (3 feet: one fully stretched arm’s length) distance between two beds for Nipah patients.
- Avoid unnecessary contact with suspected Nipah cases or use barrier nursing.
- Any spillage of body fluids in the OPD/Ward should be managed as per Infection control guidelines.
- Mortuary staff should wear PPE while handling corpse of Nipah. Air sealed bag should be used for transportation of the dead body.
______
Public health education:
In countries like Bangladesh where Nipah virus is endemic, authorities stress the importance of public awareness. An explicit warning has been made by the Health Minister A.F.M. Ruhal Haque: “Only by stopping the consumption of the raw sap, can this disease be stopped. Despite our many attempts at raising awareness, people are ignoring the warnings and as a result, are getting infected”, underlining the importance of providing information and the difficulties encountered to obtain behavior changes in target populations. In the absence of a vaccine, the only way to reduce the risk of infection in people is by raising awareness of the risk factors and educating people about the measures they can take to reduce exposure to the virus.
_
Public health educational messages should focus on:
- Reducing the risk of bat-to-human transmission: Efforts to prevent transmission should first focus on decreasing bat access to date palm sap. Freshly collected date palm juice should also be boiled and fruits should be thoroughly washed and peeled before consumption.
- Reducing the risk of human-to-human transmission: Close physical contact with Nipah virus-infected people should be avoided. Masks, gloves and protective equipment should be worn when taking care of ill people. Regular hand washing should be carried out after caring for or visiting sick people.
- Reducing the risk of animal-to-human transmission: Masks, gloves and other protective clothing should be worn while handling sick animals or their tissues, and during slaughtering and culling procedures.
_
Is India fully prepared to handle crises like Nipah?
Ebola is a disease of poverty, weak health care system, nonexistent hospital infection control, poor personal hygiene, illiteracy, fear, ignorance, superstition, and cultural practice of touching dead body. Guinea, Sierra Leone, and Liberia have all these to account for 2014 Ebola outbreak but even densely populated India has all these factors. Despite all the past information and dread coming from such large-scale morbidity, Indians are not yet at a point of effective preparedness to manage such a situation. It is not that they don’t know about the dangers of unknown, strange viruses causing large scale death and morbidity. In the case of the Nipah virus, Indians were indeed lucky that it happened in a state where the healthcare sector is better prepared than others. In any such breakout, it is important for the surveillance system to work, infected people to be isolated immediately and panic not allowed to set in. That was seen in Kerala. However, the battle is not won yet. It is only the beginning as India now needs vaccines and therapeutic drugs to fight this disease that could return anytime. While Kerala did manage to contain the impact of the virus, I worry to think how another state with less effective health systems would have dealt with a similar outbreak. In this outbreak, the tragedy was the death of the nurse who handled the early patients. While her sacrifice is immense and deserves the world’s admiration, it is also a pointer to how important it is to ensure prevention and safety of the staff dealing with it. It is indeed a wake-up call for disease control and for all neglected diseases in the country. Even as the recent Lancet report shows that India is not prepared to handle TB, this new episode once again reminds how important it is to strengthen Indian public health and disease control systems across states. In view of fruit bats in Bangladesh contaminating food sources and proved human-to-human transmission method, India cannot sit back idle even though there are not many outbreaks occurring. The NIV recently found the Nipah virus in fruit bats in West Bengal and Assam, according to a report published the Indian Journal of Medical Research. For the study, the researchers sampled 107 bats from Cooch Behar and Jaipaiguri districts in West Bengal and Dhubri in Assam, all of which are close to Bangladesh, raising the probability that the virus is circulating there. They found nine out of the 107 samples to be positive for the virus. Moreover bat population from Northeast to North-west states like Haryana has NiV antibodies means there might be active NiV infection among Indian bats too. Fruit bats from Changaroth panchayat in Kerala’s Kozhikode district are tested positive for NiV proving them to be the source of the outbreak. This indicates that there are several States in India with the virus, which means wider surveillance in animals is required. Preparedness, surveillance and constant vigil would have to be mounted continuously in the country.
_____
_____
Nipah prevention through vaccination:
Prevention through vaccination certainly includes human vaccination in endemic areas but also includes vaccinating livestock animals, particularly pigs and perhaps horses in certain endemic areas. While livestock vaccination would not prevent outbreaks of NiV in areas where it is primarily spread through date palm sap contamination, there are regions where this method could be quite effective, if it could be made cost effective. However, most efforts are being directed toward human vaccination.
The academic community of scientists has produced a number of vaccine candidates capable of complete protection against NiV disease in preclinical studies of multiple small animal and nonhuman primate models. These candidates include subunit and vectored vaccine platforms. Candidates using a vesicular stomatitis virus (VSV) vector are the most advanced having demonstrated protection in hamsters, ferrets and African green monkeys. The academic community is unlikely to be able to fund human clinical trials of these vaccines and the pharmaceutical industry traditionally has been reluctant to fund research and development of vaccines for diseases with limited incidence, albeit with devastating pandemic potential. Vaccine development requires large amounts of money. The number of people infected with Nipah is small, and so, until very recently, there has been limited investment in developing a vaccine. However, following the Ebola outbreak of 2014 it became clear that a new paradigm was needed. Thankfully, a new international coalition of governments and pharmaceutical companies called the Coalition for Epidemic Preparedness Innovations (CEPI) was formed in January 2017 to develop safe, effective and affordable vaccines for diseases with pandemic potential, such as NiV.
Recently, the WHO has declared NiV to be a priority pathogen, and CEPI is therefore focusing on NiV together with Middle East respiratory syndrome coronavirus and Lassa virus as their first disease targets. Building off previous work in the academic community, they aim to have two new experimental vaccines ready for each of these diseases beginning phase I clinical trials within 5 years so that they will be in place for efficacy studies in the event of continued or large outbreaks. It will be due to paradigm shifts, like the creation of CEPI and, hopefully, other forthcoming organizational innovations, that will finally allow for the prevention of emerging infectious diseases including NiV. CEPI has announced a grant of up to $25 million to Profectus BioSciences, Inc. and Emergent BioSolutions Inc. to develop and manufacture a vaccine against the Nipah virus.
_
Developing vaccines and antiviral therapies for Hendra and Nipah virus are also viable alternatives for mitigating disease risk. As livestock have been identified as intermediate hosts for both Hendra and Nipah virus, antiviral therapies seem less attractive given the size of horses and pigs and the significant costs associated with producing large quantities of any possible drug. Conversely, vaccination of livestock populations is a highly attractive mitigation strategy since both diseases in the target species as well as secondary transmission of virus to humans would be prevented. In areas such as Bangladesh, where no intermediate host has been definitively identified, there is a real need for the development of effective therapies and vaccine strategies to prevent infection. Similarly, for individuals who have potential occupational exposure to Hendra and Nipah virus infection, such as pig farmers and equine veterinarians, therapeutic agents and/or a vaccine to prevent infection would significantly reduce morbidity and mortality associated with Hendra and Nipah viruses.
_
Active immunization strategies:
A number of NiV vaccine studies have been conducted in which the NiV envelope glycoproteins F and G were chosen for vaccine development based on existing knowledge regarding immunity to other paramyxoviruses. There has been a variety of immunization strategies developed for HeV and NiV infection including live-recombinant virus platforms, protein subunit, virus-like particles and DNA vaccines; many of these approaches have only been examined for their ability to illicit HeV- or NiV-specific neutralizing antibody responses. Some approaches, however, have been examined for both immune response and efficacy using a variety of animal challenge models (Table below). The first vaccine approach tested utilized the attenuated vaccinia virus strain NYVAC, whereby recombinant viruses encoding either the NiV F or G glycoproteins were examined both individually and in combination to immunize hamsters. This study revealed that complete protection from NiV-mediated disease was achievable following vaccination and that an immune response to the viral envelope glycoproteins could be an important mechanism of protection. A second poxvirus-based vaccine approach was examined as a potential livestock vaccine using recombinant canarypox virus in swine. Similar to the recombinant vaccinia virus approach, the NiV F and G glycoprotein genes were inserted into canarypox virus (ALVAC) vectors and then used to immunize piglets (Table below). The ALVAC vectors expressing either NiV F or NiV G were tested individually and in combination and following vaccination, piglets were challenged intranasally with NiV. This study showed that protection from NiV-mediated illness was achievable in immunized piglets by either ALVAC vector alone or in combination, and also that vaccinated animals shed only low levels of nucleic acid detectable virus and no recoverable virus was evident. To date, the most widely evaluated henipavirus vaccine antigen has been a subunit, consisting of a recombinant soluble and oligomeric form of the G glycoprotein (sG) of Hendra virus (HeV-sG) (Bossart et al., 2005).
_
Advanced active vaccination and passive immunization platforms tested in Hendra virus and/or Nipah virus animal challenge models:
| Platform | Viral antigen target or immunogen | Animal challenge model |
| Active vaccination | ||
| Recombinant vaccinia virus | Nipah F and/or G glycoprotein | Hamstera (NiV) |
| Recombinant canarypox virus | Nipah F and/or G glycoprotein | Pigb (NiV) |
| Recombinant VSV | Nipah F and/or G glycoprotein | Ferretc (NiV), Hamsterd (NiV), nonhuman primatee (NiV) |
| Recombinant AAV | Nipah G glycoprotein | Hamsterf (NiV, HeV) |
| Recombinant measles virus | Nipah G glycoprotein | Hamster and nonhuman primateg (NiV) |
| Recombinant subunit | Hendra soluble G glycoprotein | Cath (NiV), Ferreti (HeV, NiV), nonhuman primatej (HeV, NiV) Horsek (HeV) |
| Passive immunization | ||
| Human monoclonal antibody m102.4 | Hendra / Nipah G glycoprotein | Ferretl (NiV), Nonhuman primatem (HeV, NiV) |
aHamsters immunized with NiV F and/or G glycoprotein encoding recombinant vaccinia viruses were protected against disease following intraperitoneal challenge with 103 PFU of NiV.
bPigs immunized with NiV F and/or G glycoprotein encoding recombinant canarypox viruses were protected against intranasal challenge with 2.5×105 PFU of NiV.
cFerrets immunized with NiV F and/or G glycoprotein encoding recombinant vesicular stomatitis virus (VSV) vectors were protected against lethal intranasal challenge with 5×103 PFU of NiV
dHamsters immunized with NiV F and/or G glycoprotein encoding recombinant vesicular stomatitis virus (VSV) vectors were protected against lethal intraperitoneal challenge with 105 TCID50 of NiV; or 6.8×104 TCID50 of NiV.
eAfrican green monkeys immunized with a NiV G encoding recombinant VSV vector were protected against lethal intratracheal challenge with 105 TCID50 of NiV.
fHamsters immunized with a NiV G encoding recombinant adeno-associated virus (AAV) vector were protected against lethal intraperitoneal with 104 PFU of NiV.
gHamsters and African green monkeys immunized with a NiV G encoding recombinant measles virus vector were protected against lethal intraperitoneal challenge with 103 TCID50 of NiV (hamsters) or 105 TCID50 of NiV (monkeys).
hHendra virus soluble G glycoprotein (HeV-sG) used to immunize cats protects against lethal subcutaneous (500 TCID50) or oronasal (5×104 TCID50) NiV challenge.
iHeV-sG used to immunize ferrets protects against lethal oronasal challenge with 5×103 TCID50 of HeV or 5×103 TCID50 of NiV challenge.
jHeV-sG used to immunize African green monkeys protects against lethal intratracheal challenge with 105 TCID50 of NiV or 5×105 PFU of HeV.
kHeV-sG used to immunize horses protects against lethal oronasal challenge with 2×106 TCID50 of HeV.
lA NiV and HeV cross-reactive G glycoprotein specific neutralizing human mAb (m102.4) protects ferrets against lethal oronasal challenge with 5×103 TCID50 of NiV or 5×103 TCID50 of HeV by post-exposure infusion.
mHuman mAb m102.4 protects African green monkeys by post-exposure infusion following lethal intratracheal challenge with 4×105 TCID50 of HeV or lethal intratracheal challenge with 5×105 PFU of NiV.
_____
HeV-sG vaccine:
The HeV-sG subunit vaccine is a secreted version of the molecule in which the transmembrane and cytoplasmic tail domains have been deleted from the coding sequence. HeV-sG is produced in mammalian cell culture expression systems and is properly N-linked glycosylated and retains many native characteristics including its oligomerization into dimers and tetramers, ability to bind ephrin receptors and elicit potent cross-reactive (Hendra and Nipah virus) neutralizing antibody responses (reviewed in (Broder et al., 2012)) [Table below]
_
Evaluation of the protective efficacy of henipavirus sG vaccines against lethal henipavirus challenge in laboratory animals:
| Virus | Animal model | Experimental design and results | Reference |
|
Hendra |
Ferret | Hendra-sG used to immunize followed by 10-fold lethal oronasal virus challenge | (Pallister et al., 2011b) |
| AGM (African Green Monkeys) | Hendra-sG used to immunize followed by 10-fold lethal intratracheal virus challenge | (Bossart et al., 2012) | |
| Horse | Hendra-sG is used to immunize horses followed by lethal oronasal virus challenge | (Balzer, 2011) (Middleton D., et al., submitted) | |
| Nipah | Cat | Hendra-sG or Nipah-sG used to immunize followed by 10-fold lethal subcutaneous virus challenge; Hendra-sG used to immunize followed by 100-fold lethal oronasal virus challenge | (McEachern et al., 2008; Mungall et al., 2006) |
| Ferret | Hendra-sG used to immunize followed by 10-fold lethal oronasal virus challenge | (Pallister et ell, 2011) | |
| AGM | Hendra-sG used to immunize followed by 10-fold lethal intratracheal virus challenge | (Geisbert T., 2012) | |
All studies to date have reported that all Hendra-sG immunized animals can be completely protected from infection and disease following either a Hendra or Nipah virus challenge. No evidence of virus replication or shedding has been reported in the majority of challenged subjects.
Studies showing the HeV-sG subunit immunogen as a successful vaccine against lethal Hendra virus or Nipah virus challenge have been carried out in the cat (McEachern et al., 2008; Mungall et al., 2006), ferret (Pallister et al., 2011b) and nonhuman primates (Bossart et al., 2012), and details of the results from these studies have been reviewed elsewhere (Broder et al., 2012). The success of the HeV-sG vaccine-mediated protection observed in multiple animal challenge models led to the consideration of the HeV-sG as a safe and effective vaccine for horses against Hendra virus infection in Australia following a human fatality in 2009 and the human exposure cases in 2010. The adopted equine vaccination strategy was to both prevent infection in horses and thus ameliorate the risk of Hendra virus transmission to people. A series of horse HeV-sG vaccination and Hendra virus challenge studies have been carried out in Australia; at the high containment biological safety level 4 (BSL-4) facilities of the Animal Health Laboratories (AAHL), Commonwealth Scientific and Industrial Research Organisation (CSIRO), in Geelong. The development of HeV-sG as an equine vaccine against Hendra virus was a collaborative research program between the Uniformed Services University of the Health Sciences, the Henry M. Jackson Foundation, the AAHL and Pfizer Animal Health (now Zoetis, Inc.). Findings from these initial studies were reported at Australian Veterinary Association, Annual Conference in Adelaide, in May 2011 (Balzer, 2011). The HeV-sG subunit glycoprotein was used to vaccinate horses (a 2 dose regime with a 3 week interval) and both a high and a low dose of HeV-sG antigen was examined. Following a high dose oronasal challenge with Hendra virus, all vaccinated horses remained clinically disease-free, and there was no evidence of virus replication or virus shedding in any of the immunized horses. On November 1, 2012, the vaccine called Equivac HeV® was released for use in Australia, and it is the first vaccine licensed and commercially deployed against a BSL-4 agent and currently is the only licensed prophylactic treatment for henipaviruses.
_
Cross-protection:
It is likely that novel henipaviruses induce NiV cross-reactive antibodies. There is good cross-protection provided by the G protein of HeV. Passive administration of monoclonal antibodies to HeV G protein has prevented disease in ferrets challenged with a lethal dose of NiV. Vaccinations using HeV soluble G protein have been found to protect against challenge with live NiV.
______
Animal models of henipavirus infection employed in vaccine development and efficacy studies:
Approval of human vaccines requires that the vaccine candidate is found to be efficacious in at least two animal models representing a human disease, preferably including a nonhuman primate model, while for veterinary vaccines, the requirement is efficacy in the target species. However, vaccine efficacy testing for large animals under biosafety level 4 conditions is extremely difficult and constrained. Consequently, the two animal models rule was considered when licensing the HeV vaccine for horses (Equivac HeV®) in Australia, based on the outcomes of protection studies in ferrets against HeV, and cats, ferrets, and nonhuman primates against NiV. In addition to horses, guinea pigs and ferrets were used in parallel to complement the efficacy testing.
_
Summary of vaccine efficacy reports:
Reports on vaccine candidates tested in animals for their efficacy against NiV or HeV challenge are summarized in Table below.
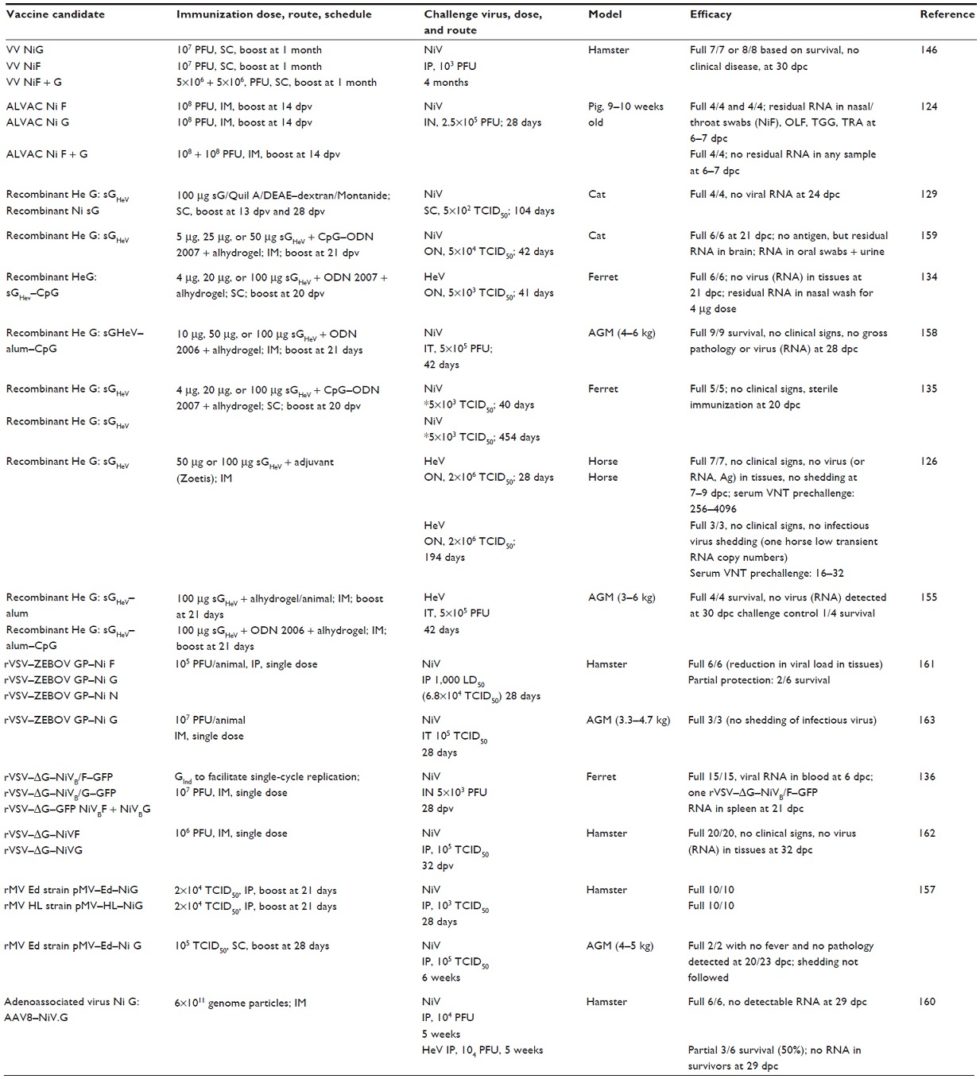
As many of them can be considered for both human and veterinary vaccines, there is no distinction made in Table above based on their intended use.
Notes:
Administration/inoculation route: IT, SC, IP, IN, and ON. *Not given. Challenge day is always in relation to the first vaccination. End, provided time of tissue collection. Protection given as “Full 6/6” indicates six animals out of six survived. For references, please see DOI https://doi.org/10.2147/VDT.S86482
Abbreviations in the table above:
VV, vaccinia virus; PFU, plaque forming unit; SC, subcutaneous; NiV, Nipah virus; IP, intraperitoneal; dpc, days postchallenge; IM, intramuscular; dpv, days postvaccination; IN, intranasal; NiF, NiV F protein; NiG, NiV G protein; OLF, olfactory bulb; TGG, trigeminal ganglion; TRA, trachea; DEAE, diethylaminoethyl; TCID50, median tissue culture infective dose; ODN, oligodeoxynucleotide; AGM, African green monkey; HeV, Hendra virus; IT, intratracheal; ON, oronasal; VNT, virus neutralization titer; LD50, median lethal dose.
_
An important strategy, when considering the protection of human population against zoonotic viruses, is interruption of the transmission cycle at the level of the intermediate host, or natural reservoir, if practical. Abolition or a very significant reduction of shedding following the challenge of vaccinated animals is considered a critical requirement for the efficacy of veterinary vaccines. The advantage of the vaccination of livestock or pets is that the vaccination also protects them. The second advantage of this approach is that the licensing of veterinary vaccines can generally take less time than the licensing of human vaccines. Licensing of the Hendra vaccine for horses (Equivac HeV®; Zoetis, Inc., Florham Park, NJ, USA) in Australia in November of 2012 is the best example. Following the fatal human case of HeV infection, Australian health authorities decided in 2009 to pursue rapid licensing of an equine vaccine for HeV. It took a total of 2 years from the decision to licensing. The successful candidate, soluble HeV G protein (HeV-sG), which was first tested in cats in 2006, was shown to be protective against HeV in ferrets prior to testing in horses. Following the successful protection experiments in AGMs against both HeV and NiV, HeV-sG may be potentially considered as a vaccine candidate for humans in the future. Even though the AGM model is not 100% lethal, additional correlates of protection and infection can be employed to assess protection, as is currently being considered for swine NiV efficacy tests, where a lack of viral RNA presence in tissues – and, importantly, a lack of shedding postchallenge – as well as cellular and humoral immune response are considered. Very interestingly, vaccination with HeV-sG protein was found to be protective against NiV in number of species, but adenoassociated virus vector expressing the NiV G protein yielded only partial protection against HeV challenge, although it was fully protective against the NiV challenge. An adjuvant suitable for the target species has to be part of the vaccine formulation for the HeV-sG based vaccine candidates to offer full protection: shedding of viral RNA was detected in cats, but not in other tested species vaccinated with HeV-sG and challenged with NiV.
_______
A VLP-based vaccine provides complete protection against Nipah virus challenge following multiple-dose or single-dose vaccination schedules in a hamster model, a 2017 study:
In this study, authors produced mammalian cell-derived native Nipah virus-like particles (VLP) composed of Nipah virus G, F and M proteins for use as a novel Nipah virus vaccine. Previous studies demonstrated that the virus-like particles were structurally similar to authentic virus, functionally assembled and immunoreactive. In the studies reported here, purified Nipah virus-like particles were utilized either alone or with adjuvant to vaccinate golden Syrian hamsters with either three-dose or one-dose vaccination regimens followed by virus challenge. These studies found that Nipah virus-like particle immunization of hamsters induced significant neutralizing antibody titers and provided complete protection to all vaccinated animals following either single or three-dose vaccine schedules. These studies prove the feasibility of a virus-like particle-based vaccine for protection against Nipah virus infection. In the studies presented here, a non-replicating VLP-based vaccine was tested to determine if it stimulated immunity and protected hamsters against direct NiV challenge. These studies demonstrated that animals vaccinated with either a three-dose series or with a single NiV-VLP dose in the context of adjuvant developed a robust NiV-specific antibody response and were fully protected. These studies provide support for further development of a VLP-based vaccine for protection against NiV infection. These studies demonstrate that the NiV-VLP vaccine has significant potential for additional development towards prophylactic use in humans. While the hamster model for NiV is not uniformly lethal, the development of significant neutralizing antibody titers in this small animal model provides a strong indicator of success. Unlike live-attenuated or live-recombinant virus platforms, there is no risk of reversion or significant secondary effects that can be seen in recombinant platforms. Unlike subunit vaccines, VLPs can express multiple viral proteins that may be invaluable for inducing protective immunity. The VLPs also present viral surface proteins in their native state allowing the VLPs to stimulate production of appropriately reactive antibodies and to bind to and fuse with cells to introduce viral antigen to major histocompatibility complex (MHC) processing. VLPs can be produced at a reasonably large scale without the need for biocontainment laboratories. VLPs could also be lyophilized to potentially reduce the need for a cold chain in resource poor environments. While additional pre-clinical testing is necessary, the NiV-VLP platform holds promise as an effective prophylactic for the prevention of NiV infection in humans.
___
Status of vaccine research and development of vaccines for Nipah virus, a 2016 study:
Despite the potential importance of NiV as an emerging disease pathogen, no specific therapeutics or vaccines are currently approved for use in humans. Current prevention of disease in endemic countries relies on behavior modifications to prevent spillover, including farming practices which decrease livestock exposure to bats, the use of bamboo skirts to prevent date palm sap contamination, and pasteurizing date palm sap. Though these approaches appear to be effective strategies of prevention, their implementation is often problematic due to cultural factors. Current efforts to develop a NiV vaccine face potential hurdles like biological feasibility, technical and regulatory assessment of candidate vaccines and financing.
Key points:
- No therapeutics or vaccines are currently approved for use in humans against NiV.
- Approximately two billion people live in areas inhabited by the Pteropus bat reservoir.
- Soluble glycoprotein vaccines and virus vector platforms are protective in experimental animals.
- Exact correlates of protection against NiV have not been completely defined.
- The sporadic nature of NiV outbreaks makes large scale Phase III clinical trials difficult to plan.
_______
_______
Nipah virus should be at the top of the list of major concerns for the human race for several reasons:
- Increased human to human transmission in recent outbreaks: Transmission of NiV has now become a concern in many hospitals in Southeast Asia. There have been several cases of doctors becoming infected from treating patients as well as infections resulting from contact with corpses. The more recent outbreaks in Bangladesh and India are suggested to have an estimated 75% or more of the known infections resulting from human to human transmission. The main mode of transmission from human to human is hypothesized to be through respiratory secretions and close contact. A mutated strain of NiV has the potential to be extremely detrimental in densely populated cities.
- Rising fatality rate in humans: The recent outbreak in Bangladesh in February of 2013 resulted in increasing the overall fatality rate of Nipah virus infection in Bangladesh to 77%. Case fatality rate in 2018 outbreak in India is 89%.
- Lack of knowledge of molecular mechanisms of infection: The molecular mechanisms of how the virus is passed from species to species are still fairly unknown.
- Shared habitats: With a rapidly growing human population in the world, particularly in Southeast Asia, there is an increase of overlapping of habitats between humans, pteropus fruit bats, and pigs. This only increases the chance of transmission of NiV between the species as well as the risk of more outbreaks.
- Pteropus fruit bat migration and pig dependence: Pteropus fruit bats are migratory animals which can survive in a wide range of environment, while much of rural Southeast Asia is dependent on their pig farms as a source of income and food. The combination of these two aspects opens up many pathways to many new countries and new populations of humans with no previous exposure to NiV.
- NiV is an RNA virus and a zoonotic virus: RNA viruses have a high mutation rate which enables them to keep a leg up on both vaccines and host immune systems. Zoonotic viruses also have a high mutation rate. Because NiV is both of these, it is hypothesized it has an extremely high rate of mutation.
______
Lessons to be learned from Nipah experience:
Ling published the editorial after Nipah outbreak in Singapore, in which he has stated that for the prevention of NiV and other zoonoses, there is a need to inspect all the imported livestock at the point of origin and when they arrive. All the facilities for their slaughter are maintained at the highest level of hygiene possible. It is also essential to take proper precautions for workers. For those who come down with these zoonoses, we must ensure that facilities for the care of patients with high level (dangerous) organisms are available, that all health care and veterinary care personnel have adequate protection, and that laboratories are equipped with BSL-3 or BSL-4 facilities to handle these dangerous organisms. A breakdown or neglect in the use of safety precautions because one is not aware that one could be exposed to dangerous organisms can lead to an outbreak of tragic consequences. After Bangladesh and Indian Nipah experience, Mackenzie in his presentation from the Global Outbreak Alert and Response Network (GOARN) platform, underlined the importance of communication between medical and veterinary health, and of the ‘One Health’ approach. He also warned about the need of improved communication between human and animal health in many countries. The key to controlling the outbreak and reducing mortalities is early detection of the outbreak and installing preventive measures as soon as possible. There is a need to have active inter-institutional and international coordination among human-animal virologists as well as virologists and ecologists to fully understand how and when the bats excrete the virus. Simultaneously there is also a need for educating the common people about personal and food hygiene. There is a need to have active inter-institutional and international coordination among human-animal virologists as well as virologists and ecologists to fully understand how and when the bats excrete the virus. Simultaneously there is a need for educating the common people about personal and food hygiene.
______
Economic and social impacts of the outbreak:
The outbreak had a devastating impact on the pig industry in Malaysia. Most of the 265 human encephalitis cases and the 105 fatalities were pig industry people, and their loss is keenly felt by all associated with the industry. Major economic costs have been incurred in controlling the outbreak, in lost domestic and export markets, and in allied businesses. The government paid US$35 million in compensation for the 1.1 million pigs destroyed at an average price of US$32 per pig. An estimated cost of US$136 million was spent in the control programme from the Department of Veterinary Services. Tax revenue estimated at US$105 million was lost from the pig industry. Approximately 618 homes and 111 shops, as well as schools and banks, were evacuated in bringing the outbreak under control, causing great financial loss to the families and business involved. In addition, the pig industry in Malaysia also provided employment to farm workers and primary supporting services like drug and vaccine sales, feed and transport. It was estimated that 36,000 people from this group had suffered from the loss of employment due to closure of farms. Also, prior to the epidemic, Malaysia had been exporting pigs to Singapore and Hong Kong. The loss of this export trade meant a loss of about US$120 million in 1999, assuming average price per pig of US$120. In addition, local pork consumption during the peak of the outbreak dropped by 80 percent and farmers supplying this market suffered financial loss estimated to be about US$124 million during the outbreak period alone. The episode caused a drastic change in the direction of the future of the pig industry in Malaysia. Pig farming is now allowed only in “identified pig farming areas”, with farmers in other areas encouraged to undertake other agricultural and livestock activities.
_
India`s Fruit exports under threat due to Nipah Virus:
India is the largest producer of bananas (26.04%), papayas (44.51%) and mangoes (40.75%). According to Agricultural & Processed Food Products Export Development Authority (APEDA), in 2016-17, India exported fruits worth of Rs 44,480 million, out of which mangoes, walnuts, grapes, bananas and pomegranates accounted for most of it. The outbreak of Nipah virus, which has resulted in the death of 17 people in Kerala, is also likely to have an adverse impact on India’s fruit exports. The virus, which is released through bats’ saliva, urine and excreta, typically spreads due to bats consuming fruits on trees. Specifically, it is the Greater Indian Fruit Bat, found abundantly across South Asia, which carries the Nipah virus. Since these bats feed on any fruit they can find, there’s a risk of exports of many fruits getting affected.
_
Church in Kozhikode to stop giving Holy Communion on tongue:
In view of the Nipah virus outbreak claiming 17 lives in Kerala, the Syro-Malabar church here has decided to stop serving the Holy Communion to the faithful on their tongue. The Thamarassery diocese here in a circular informed people about the church’s decision that the holy communion, the sacrament commemorating the last supper in which bread and wine are placed on their tongues, will henceforth be kept on their palms as a precautionary step, until further notice. Baptisms, housewarmings, marriages and prayer gatherings, which can be postponed, should be re-scheduled considering the special circumstances in the wake of Nipah, the Bishop Mar Remigiose Inchananiyil said in the circular. Believers need to be alert against the virus and strictly follow the directives issued by the state authorities to maintain extreme caution in the wake of the virus claiming 17 lives so far, he said. Unnecessary journeys, public meetings and celebrations have to be avoided totally, it said. Twenty positive cases of Nipah virus have been reported in Kerala since its outbreak in May, of whom 17 have died.
________
Deforestation and emerging diseases:
The emergence of henipaviruses has been linked to increased contact between bats and humans, paralleling the emergence of other zoonotic viruses such as SARS, coronavirus, Australian bat lyssavirus, Manangle virus, and probably Ebola and Marburg. Loss of habitat and food availability has driven bats toward human-populated areas, and encroachment by human agriculture into bat habitats creates exposure to these emerging pathogens. In the late 1990s a deadly new disease emerged from the tropical forests of Malaysia, spread by fruit bats whose natural habitat had been destroyed by deforestation. The Malaysian government was unprepared for this new disease and subsequently bore high costs from the outbreak, including more than 100 human lives lost as well as an economically devastating collapse of its pig-farming industry. Eventually, the new scourge was identified and named: the Nipah virus.
Forests play a well-known, important role in protecting biodiversity and absorbing carbon dioxide emissions. A less-recognized fact is that maintaining forests can also protect human health by helping prevent the emergence of infectious diseases. A number of infectious diseases are associated with deforestation, including yellow fever, dengue, Rocky Mountain spotted fever, and Lyme disease. Many of the diseases linked to forest loss are transmitted by insect vectors (mosquitoes that spread yellow fever, for example), but some are spread by direct contact. The emergence of the Nipah virus is a compelling example of how deforestation can contribute to the appearance of a deadly new disease.
_
In 1997, approximately 5 million hectares of tropical forest in Malaysia were slashed and burned to make room for pig farms, and the widespread burning caused a severe haze that blanketed much of the region. The haze coincided with a drought — a combination of factors believed to have reduced the number of flowering and fruiting trees that the local fruit bat population relied on for survival. With their food sources diminished, the bats sought nourishment in orchards near the pig farms; the pigs in turn ate fruit contaminated with bat urine and saliva, transmitting the virus to the livestock. The sick pigs developed respiratory illness and neurological abnormalities, and the densely packed pig farms likely contributed to the rapid pig-to-pig transmission of the virus. Farm workers subsequently experienced fever, nausea, vomiting, headache, and dizziness, and more than half of them also developed neurological abnormalities such as reduced consciousness, flaccid muscles, and spasms. Most of the victims had direct contact with the pigs. Health authorities initially believed that they were dealing with an outbreak of Japanese encephalitis, a mosquito-borne disease that is a leading cause of viral encephalitis in Asia. Yet mosquito-control measures proved ineffective at containing the disease, and autopsy and epidemiologic findings were inconsistent with Japanese encephalitis. Furthermore, when farmers sold their sick pigs across Malaysia, the disease spread — indicating that livestock, not mosquitoes, were responsible for the disease transmission. By early 1999, the disease had reached Singapore from peninsular Malaysia. It wasn’t until March 1999 — when a scientist at the University of Malaya identified the Nipah virus, a previously unknown pathogen — that the crisis began to come under control. Singapore banned the import of pigs from Malaysia and shuttered its slaughterhouses, and Malaysia took the drastic but effective step of destroying more than 1 million pigs. But by then, the virus had killed 105 people in Malaysia and sickened 160 others; the mortality rate was almost 40 percent.
_
Acknowledge the link between deforestation and disease. More than 10 years after the original outbreak, there is still no cure or vaccine for the Nipah virus. Many of the first survivors continue to suffer from chronic neurological problems. The disease is now known to also be transmittable from human to human, and it has spread to new locations. Since the first Malaysian outbreak, there have been a dozen Nipah virus outbreaks around South Asia. The most recent outbreak occurred in India, and killed 17 people so far. New infectious diseases emerging from forests have the power to devastate economies, as the Nipah virus demonstrates. When factoring the consequences of deforestation, environmentalists, policy makers, and others would do well to remember the many costs — including to human health and to the economy — associated with emerging infectious diseases. Preserving the world’s forests protects not just biodiversity, but also human lives.
_
The natural habitat for Nipah-carrying Pteropus bats is tropical forests. As these forests have been converted into agricultural lands, the bats have sought out other sources of food. In Bangladesh, the virus moves from bats to people because the bats are licking fresh date palm sap and so passing their saliva — which occasionally is infected with Nipah virus — on to people who drink the sap. Because of habitat loss, Pteropus bats in Australia are more likely to stay in suburbs where fruit trees are available, and people and horses are nearby. The bats have halted much of their annual migration because of habitat loss. As per the WHO, “There is strong evidence that emergence of bat-related viral infection communicable to humans and animals has been attributed to the loss of natural habitats of bats. As the flying fox habitat is destroyed by human activity the bats get stressed and hungry, their immune system gets weaker, their virus load goes up and a lot of virus spills out in their urine and saliva.” This means that ultimately, what is at blame is human activity only. According to Nameer, the bats can be stressed out due to various reasons, ‘It can be loss of habitat, lack of food, climate change or deforestation that can lead to the virus spilling out’.
_
Ecologic changes, human demographics, and behavior patterns such as international travel, technology and industry, and microbial adaptation have all been factors that are thought to play a role in infectious disease emergence (Morse, 1995). For the henipaviruses, speculation has focused on environmental changes such as deforestation and hunting, which subsequently affected the roosting habitats of flying foxes and placed them in closer proximity to humans (Field et al., 2001; Daszak et al., 2004; Breed et al., 2005). It should be noted also that in Bangladesh, located at a more subtropical latitude, all Nipah outbreaks occurred in the first half of the year, suggesting that perhaps climate change or bat activities affected by seasonal change may also have played a role. Until a better understanding of the causes of these newly emergent viruses is obtained, future outbreaks are likely to recur. Research is continuing to identify the factors that have led to the emergence of Hendra and Nipah viruses in domestic animal and human populations.
_
Reducing the impact of deforestation is an important measure in Nipah Virus prevention. The flying fox bat loves to feed in the trees. Planting trees restore this important feeding ground and reduce contact between the bats and the horses/pigs. Nutritional stress leads to relocation of bat populations that may then lead to Nipah Virus outbreaks. Trees planted in reforestation initiatives include the fruit trees that the bats prefer. They reduce nutritional stress and provide excellent buffer zones between areas where people are settled and bat populations.
______
______
Nipah Virus and the Potential for Bioterrorism:
Nipah is just one of many viruses that are available to terrorist groups for development as a bioweapon. In 1999 this virus was first found and noted to be very easily disseminated to humans through inhalation and ingestion. Even though there are many potential pathogens available, the Nipah virus has proven itself to be one of the most dangerous and advantageous. While no country is known to have researched its weaponization, its potential for widespread use and 50 percent mortality rate make it a bioweapon to watch for.
The Centers for Disease Control and Prevention (CDC) has listed the Nipah virus as a critical biological agent, Category C. Category C agents are emerging pathogens that could be engineered for mass dissemination in the future because of:
-Availability
-Ease of production and dissemination
-Potential for high morbidity and mortality rates and major health impact
With this categorization the virus is a living pathogen that can be developed as a bioweapon with the right knowledge, and equipment. For the virus to be weaponized it needs to be purified, stabilized and properly sized. Since this is a living virus the bioterrorist agent can be replicated once disseminated (Ryan & Glarum, 2008). At this time, there is no information about how this virus could be manufactured to become a bioterrorist agent, but with the right knowledge the potential is there.

_
According to Kortepeter and Parker (Kortepeter & Parker, 1999), for a biological agent to be used for a greatest plausible occurrence, an agent must have specific properties:
- the agent should be highly lethal and easily produced in large quantities
- Given that the aerosol route is the most likely for a large-scale attack, stability in aerosol and capability to be dispersed (17,000 to 5,000 nanometers [nm] particle size) are necessary
- being communicable from person-to-person, and
- having no treatment or vaccine
In using the above criteria the Nipah virus would make a credible biological threat for a domestic or international terrorist group. Host bats being plentiful in Australia and southern Asia would make it easy to obtain the saliva, feces or urine of these hosts for initial development of the virus. The Nipah virus being “150 to 200 nm in diameter and 2000 nm long”, it could be used in an aerosol form for dispersement. According to Gurley et al., there is significant evidence that there is person-to-person communicability and according to the WHO, “there are currently no drugs or vaccines available to treat Nipah virus infection. Intensive supportive care with treatment of symptoms is the main approach to managing the infection in people”.
_
The Nipah virus should be a concern for any government as a potential for a bioterrorist attack. As with the 9/11 and the anthrax attacks in 2001 there could be significant fear, panic, economic issues and social disruption if this virus was used. With a mortality rate of 40% to 100% (Lam, 2002; WHO, 2009), and an incubation period of up to 45 days (WHO, 2009), this could definitely be a pathogen of choice for terrorists. The ease of access to the virus itself from fruit bats, to pigs and to humans, not to mention the transmissibility ease through inhalation and ingestion, makes this the perfect biological weapon. The disease this virus manifests, from flu type symptoms to severe encephalitis, will cause significant fear to the public and will stress healthcare facilities if a large outbreak occurs. This virus also has no known cure as of this date, even though there are developments in that direction. The Nipah virus needs to continue to be monitored and treatment options along with vaccine development needs to be continuous until this threat is diminished.
______
______
Public information:
What is Nipah?
Nipah is a disease caused by a virus which commonly affects animals like bats, pigs, horses, etc. The virus can spread from animals to humans and can cause serious illness among humans. Bats are the reservoir of virus, without getting disease from virus.
How does it spread?
Spread of Nipah virus to humans may occur after close contact with other Nipah infected people, infected bats, or infected pigs. Bat secretions and excretions laden with virus can infect people during fruit tree climbing, eating/handling contaminated fallen fruits or consuming raw date palm sap/juice or toddy. Nipah can spread from infected pig to humans via close contact and respiratory secretions. Human to Human infection can occur from close contact with persons affected with Nipah at home while providing care and in hospital setting if appropriate measures are not taken. Handling of dead bodies suspected of death due to Nipah virus should be done in accordance with the government advisory. During this emotional moment, traditional rituals and practices may need to be modified to prevent the exposure of family members to the disease.
Who are at high risk of developing Nipah virus infection?
-People who are exposed to areas inhabited by fruit bats/ articles contaminated by secretions such as, unused wells, caves, fruit orchards, palm sap etc. are likely to be at higher risk of infection
-Persons with direct contact with sick pigs or their contaminated tissues
-Persons in close contact with living Nipah patient at home/hospital or deceased Nipah patient during burial or cremation rituals
-Health care workers having direct contact with probable or confirmed cases without using standard precautionary measures
What measures of prevention should be taken in high risk areas?
-Wash hands with soap and water after coming in contact with a sick person or animal
-Avoid consuming raw date palm sap or toddy
-Consume only washed fruits
-Avoid consuming half eaten fruits from the ground
-Handling of dead bodies should be done in accordance with the government advisory
What is the common presentation of the Nipah Virus Disease?
The illness commonly presents as brain fever. In some cases during early part of illness it may also present with fever with persistent cough and difficulty in breathing.
____

_____
If you live in a region which is on an alert due to NiV, you can help yourselves in the following ways:
Animal-to-human transmission prevention:
-Do not consume date palm sap as the chances of it being contaminated are very high. Fruit bats are very likely to visit the date palm trees and lick the sap.
-Domestic animals can also be the carriers of NiV as fruit bats often drop partially eaten fruits which can be consumed by them. Try to keep them indoors and feed them yourself or if there’s a chance of them being infected, keep distance from them and get them treated.
-Don’t climb trees where bats may have left their saliva or secretions.
Human-to-human transmission prevention:
Epidemiological surveys have suggested that human-to-human transmission of NiV is not very common. However, it is possible to get infected with NiV if one gets in touch with the secretions and excretions of infected patients. The primary carriers of NiV in humans are the respiratory secretions. If you are near anyone who has the infection and have the chance to breathe in their out-breath, you can get infected too. The risk gets very high is the patient has respiratory symptoms, such as coughing and sneezing. Because of the same reason, never share food, bed or be very close to a sneezing or coughing NiV patient. At any cost, one should not come in contact with the saliva of an infected person. Traces of NiV are present in urine as well, besides saliva and throat swabs. So, be cautious if you are sharing a washroom with an infected person.
______
______
Moral of the story:
_
- Nipah is a viral zoonotic disease caused by nipah virus (NiV) of the Henipavirus genus of Paramyxoviridae family, which can cause rapid, fatal respiratory and neurologic disease in both humans and animals. Pteropus bats (fruit eating bat species, popularly known as flying foxes) are reservoir host of the virus. Antibodies to Nipah virus have been found in 9–25% of pteropus bats in Malaysia, Cambodia, Thailand, and Bangladesh; although it is difficult to find a bat carrying the Nipah virus. Experimental infection of Pteropus bats with NiV does not cause illness in the bat and the virus cannot be found in experimentally infected bats after a few weeks. The time window in which the bats are capable of passing on the infection to other animals or humans is quite small.
_
- Nipah virus outbreaks have been reported in Malaysia, Singapore, Bangladesh and India. From 1998 to May 2018, there have been at least 608 cases of NiV human infections with caused at least 365 human deaths in the world. This is a small number considering the world statistics for other infectious diseases. Although Nipah virus has caused relatively few outbreaks in Asia, it infects a wide range of animals and causes severe disease and death in people, making it a public health concern.
_
- Although Nipah cases tend to occur in a cluster, 18 % of cases in Bangladesh were isolated. A single confirmed case of NiV infection constitutes an outbreak. Further cases may be identified through active case finding among close contacts with similar exposure. Contact tracing, aggressive monitoring, quarantining of contacts, and isolating suspect/probable/confirmed Nipah cases are effective forms of infection control and containment.
_
- Geographic distribution of Henipaviruses overlaps with that of Pteropus category. This hypothesis was reinforced with the evidence of Henipavirus infection in Pteropus bats from Australia, Bangladesh, Cambodia, China, India, Indonesia, Madagascar, Malaysia, Papua New Guinea, Thailand and Timor-Leste. Despite having a relatively high infection rate, the species of Pteropus fruit bats are not susceptible to the nipah virus. Infected bats shed virus in their excretion and secretion such as saliva, urine, semen and excreta but they are symptomless carriers. Serological evidences indicate that circulation of Henipaviruses in bats is not limited to species belonging to the genus Pteropus, but also extended to a wider range of both frugivorous and insectivorous bats. Bats are natural reservoirs of many pathogens and since they are highly mobile, social, and long-lived, they can readily spread disease.
_
- Nipah (NiV) and Hendra (HeV) viruses are negative-sense, single-stranded zoonotic RNA viruses that have emerged in the Asia-Pacific region. Both are RNA viruses belonging to the Paramyxoviridae family and grouped under the genus Henipavirus, since they share antigenic, serological, and ultrastructural characteristics and differ from other paramyxoviruses. Paramyxoviridae is a family of viruses in the order Mononegavirales. Vertebrates serve as natural hosts; no known plants serve as vectors. There are currently 49 species in this family, divided among 7 genera. Diseases associated with this negative-sense single-stranded RNA virus family include measles, mumps, and respiratory tract infections.
_
- The NiV genome is a negative-sense nonsegmented RNA of 18,246 nucleotides (Malaysian isolate) or 18,252 nucleotides (Bangladesh isolate). [18,234 nucleotides for HeV] Like other paramyxoviruses, the genome contains six transcriptional units encoding six structural proteins: nucleocapsid (N), phosphoprotein (P), matrix protein (M), fusion protein (F), receptor-binding (G), and polymerase (L), and three nonstructural proteins, C, V, and W. Similar to all paramyxoviruses NiV RNA replication occurs in the cyptoplasm. All but the P gene are monocystronic, in that they code for a single protein. The P gene also encoded for the C, V, and W proteins which play a role in the virulence of NiV by evading interferon. The V and W proteins are major determinant of nipah pathogenesis and thereby determine nipah disease course.
_
- Phylogenetic analysis demonstrated that two phylogenetic lineages were formed for NiV sequences, one including Bangladesh and India sequences, and the other Malaysia and Cambodia sequences. By phylogenetic analysis it was unmistakable confirmed that the same NiV strain circulates in India and Bangladesh, and that it was different from that circulating in Malaysia and Cambodia.
_
- RNA viruses have a high mutation rate which enables them advantage over both vaccines and host immune systems. Zoonotic viruses also have a high mutation rate. Since NiV is both an RNA virus and a zoonotic virus, it has high rate of mutation. The rate at which mutations fix in a population is termed the evolutionary rate, which is measured by comparing the genomes of different isolates of a virus collected at several different time points. In most viruses evolutionary rates correlate well with mutation rates. The mean value of Nipah virus N gene evolutionary rate is 6.5 × 10−4 substitution per site per year. Influenza A virus evolutionary rate ranged from a minimum of 1.43 × 10−3 substitution per site per year (H3N8) to a maximum of 11.62 × 10−3 (H7N7).
_
- Hendra and Nipah viruses possess an exceptionally broad species tropism; and except pteropid bats, both natural & experimental infections have demonstrated their capacity to cause disease which can often be fatal in horses, pigs, cats, dogs, ferrets, hamsters, guinea pigs, monkeys, humans and chicken embryos spanning 7 Orders (6 mammalian and one avian). The broad species tropisms and the ability to cause fatal disease in both animals and humans distinguish HeV and NiV from all other known paramyxoviruses. NiV can be transmitted to humans by three different routes: Pteropus fruit bat to human, pig to human, and human to human.
_
- Pigs have been implicated as intermediate amplifying hosts for NiV. Horses are implicated as intermediate amplifying hosts for HeV. The intermediate host is not immune to the disease as compared to the host reservoir fruit bat. Serological studies indicated that NiV occurred in dogs, cats, horses and goats in the outbreak areas of Malaysia but all were effectively dead-end hosts. The uniformly negative serology results from surveyed peridomestic rodents, insectivores, and birds in Malaysia indicate that these animals did not play a role as secondary reservoirs for Nipah virus.
_
- The major determinant of species and cellular tropism of HeV and NiV is derived from the functions of the viral envelope glycoproteins (the receptor-binding G protein, and the fusion protein F) which are the mediators of virus attachment and host cell infection. The HeV and NiV G glycoprotein bind to the host cell membrane proteins ephrin-B2 and ephrin-B3. These glycoproteins work in concert; thus, in the case of NiV or HeV, the binding of G to a cellular receptor (ephrin B2/ephrin B3) induces a conformational cascade in G that ultimately triggers F to execute virus-cell or cell-cell membrane fusion. Membrane fusion is essential for viral entry into host cells and for cell-cell fusion, a hallmark of the disease pathobiology. Plasma membrane fusion of infected cells with uninfected ones further facilitates virus spread throughout the host, and contributes to the pathology of henipavirus disease. The new viral cells are able to fuse together and create a huge multinucleated cell called syncytia. Ephrin B2 is important in mammalian host development and it is a highly conserved protein across mammalian species, expressed on lymphocytes, neurons, smooth muscle cells, and endothelial cells surrounding small arteries, corresponding with virus distribution in an infected host as determined by immunohistochemistry. The cellular function of ephrin B2 is to regulate processes such as neurogenesis, angiogenesis, proliferation, and remodelling, as well as immune activation and bone formation. Using ubiquitous and conserved proteins as receptors, henipaviruses not only have a wide host range, but they also infect a wide range of tissues and organs within each individual host species. Ephrin-B2 expression is prominent in arteries, arterioles and capillaries in multiple organs and tissues while ephrin-B3 is found predominantly in the nervous system and the vasculature. The susceptible host species and associated cellular tropism and pathology of Hendra and Nipah virus has in large part been explained by their use of the highly conserved ephrin-B2 and -B3 proteins as entry receptors.
_
- Nipah viruses are deadly because its use of highly conserved cell surface molecules (Ephrin) as entry receptors; and its highly effective replication and fusion strategies are important characteristics responsible for its high pathogenicity. Their unique genetic constitution, high virulence and wide host range set them apart from other paramyxoviruses. The case fatality rate is estimated to be approximately 70% but varies from 40 % to 100%. This rate can vary by outbreak depending on the severity of the disease, the availability of adequate healthcare facilities and local capabilities for epidemiological surveillance.
_
- Despite absence of infection except in dendritic cells, all leukocyte types especially lymphocytes are capable of binding NiV and transmitting it to susceptible cells. The high trafficking capability of lymphocytes and their constant interaction by rolling onto the endothelial cells could explain disseminated microvascular NiV infection in multiple organs in the absence of detectable viremia. This explains why NiV has been isolated from urine and cerebrospinal fluid from infected patients, but there is no report of isolation from serum in humans. Consequently peripheral blood mononuclear cells appear much more appropriate than serum for NiV diagnosis and virus isolation from infected patients. The lymphocyte-bound NiV particles are remarkably stable and retain infectivity for a prolonged period. I hypothesize that since lymphocytes are acting as carriers of NiV and thereby giving protection to NiV, cellular adaptive immune response plays little role in either protection by vaccination or natural immunity. On the other hand, neutralizing antibodies are highly protective when elicited/administered before or shortly after infection.
_
- Due to deforestation in Malaysia, the bats sought nourishment in orchards near the pig farms; the pigs in turn ate fruit contaminated with bat urine and saliva, transmitting the virus to the livestock. The sick pigs developed respiratory illness and neurological abnormalities, and the densely packed pig farms likely contributed to the rapid pig-to-pig transmission of the virus. Transmission between pigs occurs through direct contact, exposure to infected secretions, and contact droplet transmission. Mortality in pigs is only 5% but transmission is 100%. Nipah virus is highly contagious in pigs, which can act as amplifying hosts and shed this virus in respiratory secretions and saliva. Farm workers subsequently developed nipah encephalitis as pigs transmit NiV to people through sputum, splashing urine, and large respiratory droplets, facilitated by a characteristic barking cough in infected pigs. Pigs transmit NiV differently than bats in that they shed the virus primarily by coughing and the expulsion of respiratory secretions and saliva.
_
- While the outbreak in Malaysia had progressed from the natural host (fruit bats), to amplification host (livestock) and finally to humans, in Bangladesh no amplification host was needed. People were somehow being directly infected by fruit bats. There is epidemiological evidence that zoonotic transmission of Nipah virus during outbreaks in Bangladesh occurs mainly through the consumption of raw date palm sap. This date palm sap is harvested at night in Bangladesh during the winter months and bats have been observed drinking from the sap stream or the collection pot. It is thought that bats contaminate the date palm sap with their saliva, urine or faeces containing Nipah virus while drinking from it, and people who subsequently drink the contaminated sap can acquire Nipah virus infection. In Bangladesh, all Nipah outbreaks occurred in the first half of the year, suggesting that perhaps climate change or bat activities affected by seasonal change may also have played a role. This zoonotic transmission route was investigated experimentally in the Syrian hamster model. Five of eight Syrian hamsters exposed to a high dose of Nipah virus via drinking of artificial palm sap containing the virus indeed developed neurological signs requiring euthanasia. Drinking of virus contained palm sap indeed transmit nipah and virus replication was detected in the respiratory tract & CNS, but not in the intestinal tract.
_
- Human-to-human transmission of Nipah virus can occur, but that transmission is rather inefficient, and probably requires prolonged close contact. Among a cohort of 338 health care workers who cared for Nipah patients at 3 Malaysian hospitals and reported a combined 89 episodes of Nipah patient blood or body fluid directly contacting their bare skin, 39 splash exposures of blood or body fluid into their eyes, nose or mouth, and 12 needle stick injuries, none developed clinical illness associated with Nipah infection. This proves that close contact with nipah patient leads to human transmission and not accidental exposure. Human-to-human transmission requires intimate contact with high levels of bodily secretions (respiratory secretions, saliva, urine, etc.). The risk of wide-spread transmission is therefore low. This is also reflected in estimated R0 ~0.4 for Nipah virus. From 2001 till date, around half of reported cases in Bangladesh were due to human-to human transmission by close contact. In Siliguri, India, transmission of the virus was reported within a health-care setting, where 75% of cases occurred among hospital staff or visitors. All persons who transmitted Nipah died, suggesting that late stages of infection, presumably with high virus titers, increase the risk of transmission. Physical contact with NiV-infected patient who later died is the strongest risk factor for developing NiV infection in the outbreak. Human-to-human transmission was modelled in the Syrian hamster to determine the route through which Nipah virus is transmitted. Upon intranasal inoculation of Syrian hamsters with Nipah virus, transmission to naïve hamsters was observed only in hamsters that were in direct contact, but not via fomites or aerosols.
_
- Nipah virus transmission via respiratory secretions is the most important mode of viral transmission compared to other modes of transmission in animals and humans. Respiratory secretions appear to be particularly important for human-to- human transmission of NiV. NiV RNA is readily identified in the saliva of infected patients. Persons who are in a room where a Nipah patient is coughing or sneezing are at increased risk of Nipah virus infection. Across all recognized outbreaks in Bangladesh from 2001 through 2007, Nipah patients with respiratory symptoms were more likely to transmit Nipah. Nipah is spread by droplet contact transmission and not by airborne transmission. Droplets containing microorganisms can be generated when an infected person coughs, sneezes, or talks. Droplets are too large to be airborne for long periods of time, and quickly settle out of air. Droplet contact transmission can be reduced with the use of personal protective barriers, such as face masks (for patients and health care workers) and goggles. Frequent hand cleansing, especially with instant hand sanitizers, can help prevent droplet transmission. Maintain >1 full-stretched arm distance (1 metre or 3 feet) from patient to prevent droplet contact transmission. To prevent nosocomial transmission of Nipah virus; isolation, sanitation, barrier nursing, and use of PPE (contact and droplet precautions) seem prudent when taking care of any patient with suspected Nipah virus infection.
_
- Different patterns of transmission have been associated with recurrent outbreaks caused by Nipah virus (NiV) in Bangladesh, where person-to-person transmission is a major pathway for human infection, compared to an outbreak in Malaysia, where pig-to-human transmission accounted for virtually all human infections. Although higher levels of oral shedding of the Bangladesh strain of Nipah virus might be a key factor in onward transmission in outbreaks among humans, active transfer of infected bodily fluids consistently results in transmission, regardless of the virus strain. So a critical driver of onward human to human NiV transmission is the nature of the interaction between subjects and that its impact may be irrespective of virus strain. Human to human transmission in Bangladesh and India, and absence of human-to-human transmission in Malaysia is most likely not due to a difference in the ability of the virus to transmit, but most likely due to high use of standard and respiratory precautions in Malaysia and cultural differences resulting in more contact between sick individuals and their family members in Bangladesh. The pattern of the outbreaks in Bangladesh and India suggests that person-to-person transmission is more dependent on the characteristics of the occasional Nipah transmitter than a specific virus strain. Human-to-human transmission of NiV is more plausibly underpinned by environmental factors in play with each outbreak event, than by inherent differences in tissue tropism or pathogenicity between different NiV strains.
_
- Virus is a link between living and non-living matter. Although virus is an obligate intra-cellular parasite, virus outside living cells exist as particles which can be viable or non-viable. Non-viable virus particle in the environment is dead, cannot enter in host cell (e.g. empty capsids lacking any genetic material). Viable virus particle in environment is a state between living and non-living matter. Such particles can survive for hours to days depending on environmental conditions such as temperature, pH and chemical milieu. They can enter host cell and they are infectious or non-infectious. Infectious means completion of the replication cycle, from entry to synthesis of new particles in the host cell independently. Non-infectious viable virus particle means it is unable to replicate in host cell on its own due to variety of reasons although it can enter host cell. Non-infectious virus particle can replicate with assistance. Smallpox virus can remain infectious for long periods even when outside of a host while Ebola virus left alone on a surface will begin to disintegrate rather quickly. Airborne transmission of Chickenpox, Influenza, Measles and Smallpox proves that these viruses can survive in air for long period of time outside body.
_
- NiV can survive for up to 3 days in some fruit juices or fruit pulp, and for at least 7 days in artificial date palm sap (13% sucrose and 0.21% bovine serum albumin in water, pH 7.0) held at 22°C. This virus is reported to have a half-life of 18 hours in the urine of fruit bats.
_
- The sensitivity of henipaviruses to pH, temperature and desiccation indicates a need for close contact between hosts for transmission to occur, although under ideal conditions henipaviruses can persist for extended periods facilitating vehicle-borne transmission. Transmission of NiV through fomites is plausible. Data collected from various hospitals in Bangladesh where patients infected with NiV were treated showed a high amount of NiV RNA on hospital surfaces. To reduce the risk for fomite transmission of NiV, infection control should target hospital surfaces.
_
- Nipah virus is readily inactivated by soaps, detergents and many disinfectants. The effect of heat may depend on the substrate. Nipah viruses are rapidly inactivated by desiccation at both 22°C and 37°C. Nipah virus concentrations decreased but the virus was not completely eliminated in artificial palm sap held at 70°C for 1 hour. However, it was completely inactivated by heating at 100°C for more than 15 minutes.
_
- From various Nipah virus outbreaks in humans, the R0 (basic reproduction number) was estimated to be ~0.4. For an infection to spread through a population, R0 needs to >1. When R0 is <1, the infection will eventually die out. Nipah outbreaks have tended to fizzle out after four or five chains of transmission in humans.
_
- Although average incubation period for nipah is 4 to 20 days in humans; it can be as short as two days or as long as two months.
_
- Infectious dose and period of infectivity are unknown in nipah in humans. A study found that the minimal NiV infectious dose to infect ferrets oronasally is 5×103 TCID50. The minimal infectious dose by aerosol inoculation is 0.6–3.0 TCID50 for influenza A in humans. Obviously minimal infectious dose of nipah by droplet transmission would be much larger than influenza virus in humans. Such a high infectious dose and no airborne transmission definitely reduce human to human transmission exemplified by R0 of 0.4. Experimental infections in pigs and hamsters suggest that oro-pharyngeal virus shedding may start as early as 2-4 days after infection and persist for up to 3 weeks. So it is prudent to assume that infected human is infectious from second day of entry of nipah virus for at least 3 weeks. The infected patient is maximally contagious when patient is gravely ill. Experimental infection in hamsters showed that those who did not develop signs of disease also shed viral RNA. Thus asymptomatic infected humans can also shed virus and therefore indeed infectious.
_
- Nipah virus can cause asymptomatic infections in some patients; in the Malaysia and Singapore outbreaks between 17% and 45% of infections were asymptomatic. There has not been any evidence for asymptomatic infection in outbreaks in Bangladesh although cases of mild illness were identified.
_
- Relapse occurs in 3–8% of patients who had recovered from clinical illness, occasionally causing more severe clinical symptoms than the initial manifestation. Relapsed-encephalitis caused by Nipah virus has been reported in people from several months to as long as 11 years following infection.
_
- Late-onset encephalitis occurs in 3% of infected patients who were initially asymptomatic (who had only serological evidence of infection but not ill) or non-encephalitic, and has been reported to occur up to several months or even years after the initial infection.
_
- Approximately 20% of patients of nipah encephalitis are left with residual Neurologic sequelae including cognitive deficits, verbal impairment, cranial nerve palsy, cerebellar abnormalities, and persistent vegetative state; and Neuropsychiatric sequelae including personality changes and major depression.
_
- In the Nipah virus outbreak, the clinical and laboratory speculation initially centered on Japanese encephalitis (JE) virus as the causative agent; however, several features of the outbreak argued against this possibility. First, JE virus appeared an unlikely agent in an outbreak that affected mostly adults rather than children in an area of endemic JE virus transmission. Second, most, if not all, patients had a history of direct contact with pigs in Malaysia. Additionally, there was clustering of cases with higher infection rates in the same households than would be expected with JE. Finally, many patients had been previously vaccinated against JE with a standard vaccine and were therefore likely to have developed protective immunity against the virus. On the other hand, three cases of Japanese Encephalitis have been detected from the suspected Nipah virus infection samples that were sent to the National Institute of Virology in Pune in May 2018. So Japanese Encephalitis virus should also be considered for diagnosis while screening for Nipah virus infection.
_
- Laboratory diagnosis of nipah virus infection in suspected or probable nipah case is done by using combination of tests:
-IgM antibody against Nipah virus by ELISA in serum or cerebrospinal fluid
-Nipah virus RNA identified by RT-PCR from respiratory secretions, urine, or cerebrospinal fluid
– Isolation of Nipah virus from respiratory secretions, urine or cerebrospinal fluid (only done in BSL-4 lab).
An initial IgM anti-Nipah virus antibody response was noted in about half of patients on day one of symptoms, rising to 100% from day 3–9. However, studies have shown that some patients confirmed by RT-PCR or viral isolation of NiV may not have detectable IgM (false negative result). In surveillance programs, a positive IgM test would need to be followed with a positive virus neutralization result to confirm the diagnosis and prevent a costly and unnecessary response to a false positive result. I repeat, a single confirmed case of NiV infection constitutes an outbreak.
_
- In patients who have died, immunohistochemistry can be used to detect viral antigens in tissues. Immunohistochemistry can be performed on preserved tissues allowing a diagnosis to be made retrospectively. It also has the advantage that testing can be done in the absence of a BSL-4 facility.
_
- Most of the CT scans of the brain did not reveal any abnormality in nipah encephalitis. However, MRI abnormalities were unique, often showing multiple, small (less than 5 mm), asymmetric focal lesions in the subcortical and deep white matter without surrounding edema. The lesions were attributed to widespread microinfarctions from underlying vasculitis of cerebral small vessels. These were in stark contrast to the bilateral thalamic and basal ganglia involvement in neuroimaging of Japanese Encephalitis, and abnormal signal/patchy enhancement of the temporal lobe indicative of herpes simplex encephalitis.
_
- There is certainly a paucity of blinded, randomized, controlled trial data on the efficacy of ribavirin and acyclovir in nipah but Malaysian and Singapore outbreak studies have shown efficacy of ribavirin and acyclovir in nipah encephalitis. Recent animal studies have revealed no therapeutic benefit of either drug. One Indian nurse responded very well to treatment using Ribavirin tablets in May 2018 outbreak. Recent animal study demonstrates that favipiravir has potent antiviral activity against henipaviruses.
_
- A neutralizing human monoclonal antibody (hmAb), m102.4 recognizes the receptor binding domain of the HeV and NiV G glycoproteins. This hmAb potently neutralized both viruses in vitro and maintained its biological activity in vivo suggesting its possible utility as a passive therapeutic modality following henipavirus infection. This hmAb m102.4 is an ideal therapeutic, targeting a critical functional domain of the G glycoprotein of nipah virus, and is a fully human IgG.
_
- Viral interference may be a useful mechanism to combat Nipah infections by expression of the attachment glycoprotein on host cells.
_
- Vaccine development requires large amounts of money. The number of people infected with Nipah is small, and so until very recently, there has been limited investment in developing a vaccine. To date, the most widely evaluated henipavirus vaccine antigen has been a subunit, consisting of a recombinant soluble and oligomeric form of the G glycoprotein (sG) of Hendra virus (HeV-sG). The HeV-sG glycoprotein subunit vaccine is composed of the entire ectodomain (amino acids 76–604) of the HeV G glycoprotein. HeV-sG vaccine has been shown to provide protection against henipavirus challenge in cats, ferrets, monkeys and now horses, and has been licensed and deployed for use in Australia. Vaccinations using HeV soluble G protein have been found to protect against challenge with live NiV. Efforts are currently under way to develop HeV-sG for human use as well as for use in pigs. Very interestingly, vaccination with HeV-sG protein was found to be protective against NiV in number of species, but adenoassociated virus vector expressing the NiV G protein yielded only partial protection against HeV challenge, although it was fully protective against the NiV challenge.
_
- Forests play a well-known, important role in protecting biodiversity and absorbing carbon dioxide emissions. A less-recognized fact is that maintaining forests can also protect human health by helping prevent the emergence of infectious diseases. A number of infectious diseases are associated with deforestation, including yellow fever, dengue, Rocky Mountain spotted fever, and Lyme disease. There is strong evidence that emergence of bat-related viral infections communicable to humans and animals have been attributed to the loss of natural habitats of bats due to deforestation. As the flying fox habitat is destroyed by human activity, the bats get stressed and hungry, their immune system gets weaker, their virus load goes up and a lot of virus spills out in their urine and saliva. This means that ultimately, what is at blame for nipah is human activity only.
_
- The Nipah virus should be a concern for any government as a potential for a bioterrorist attack due to ease of access to the virus from fruit bats; mortality rate of 40% to 100%; transmissibility ease through inhalation and ingestion; and having no treatment or vaccine.
_
- NiV is a zoonotic RNA virus having exceptionally high rate of mutation; and that if a mutated human-adapted strain evolves that can be transmitted airborne to infect communities, high population densities and global interconnectedness would rapidly spread the infection worldwide. Emergence of a Nipah virus strain with more efficient person-to-person transmission (airborne and high R0) could produce a deadly global pandemic.
_______
Dr. Rajiv Desai. MD.
July 7, 2018
_______
Postscript:
Shoddy media reporting in India about Nipah outbreak only created fear and anxiety. Bad information can be worse than the virus itself.
______
52 comments on “NIPAH”
Leave a Reply

What’s Taking place i’m new to this, I stumbled upon this I have discovered It positively useful and it has helped me out loads. I’m hoping to contribute & help other customers like its helped me. Good job.
Thanks again for the blog article.Really looking forward to read more. Really Cool.